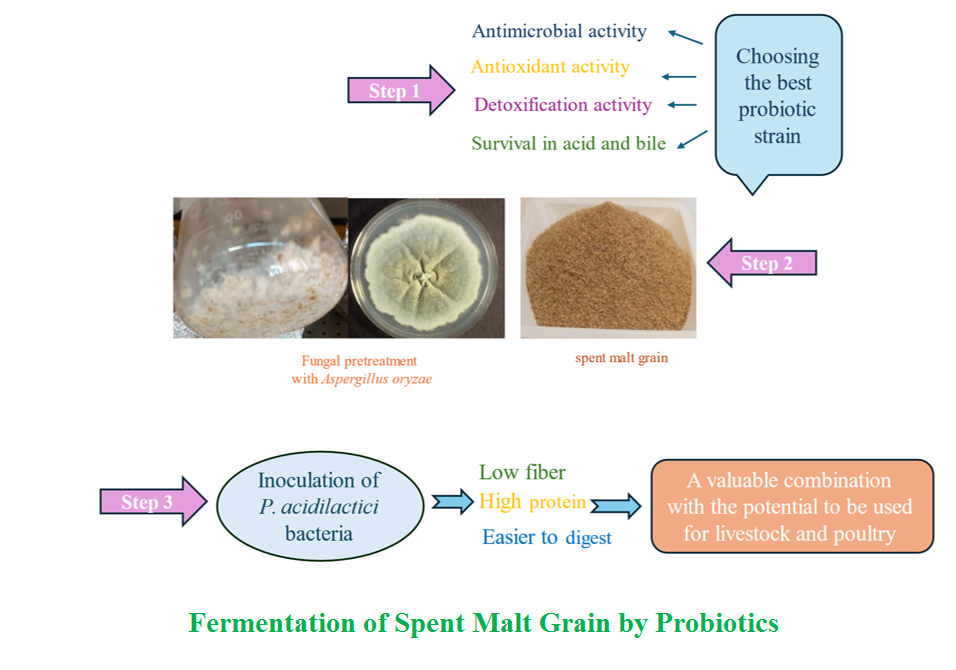
Fungal Pretreatment Strategy to Enhance Growth of Pediococcus acidilactici via Solid-state Fermentation of Spent Malt Grain
Applied Food Biotechnology,
Vol. 11 No. 1 (2024),
18 Aban 2023
,
Page e32
https://doi.org/10.22037/afb.v11i1.46281
Abstract
Background and Objective: Growing probiotic bacteria on agricultural wastes via solid-state fermentation presents challenges, compared to fungal fermentation. Spent malt grain, a byproduct of non-alcoholic malt beverage production, is rich in proteins, fibers, minerals and bioactive compounds, making it a promising substrate for solid-state fermentation. However, the lignocellulosic structure of spent malt grain limits nutrient accessibility for probiotics. Fungal pretreatment with Aspergillus oryzae can degrade these complex fibers, enhancing nutrient availability and fermentation potential of the spent malt grain. This study aimed to select probiotic bacteria with superior characteristics, assess fungal pretreatment effects on viability as well as changes in spent malt grain composition during solid-state fermentation.
Material and Methods: Four probiotic strains were assessed for antimicrobial, antitoxic and antioxidant characteristics, as well as acid and bile resistance. Pediococcus acidilactici was selected as the optimal strain. Solid-state fermentation was carried out using spent malt grain with fungal pretreatment to enhance Pediococcus acidilactici growth.
Results and Conclusion: Probiotic viability increased from 9.4 to 9.76 log CFU.g⁻¹, protein content increased by 32.45% and ash by 59.6%, while neutral and acid detergent fibers decreased by 21.6 and 9.1%, respectively. These results demonstrated effectiveness of fungal pretreatment combined with solid-state fermentation, providing a sustainable method for enhancing probiotic growth on spent malt grain with potential uses in ruminant and poultry feeds.
Conflict of interest: All authors declare no conflict of interest.
- Introduction
Solid-state fermentation (SSF) is an effective method for enhancing the nutritional value of agricultural and industrial wastes. It decreases biological wastes, prevents its accumulation in the environment and generates beneficial and nutritious compounds in primary substrates, allowing for reuse in the food cycle [1]. Brewers’ spent grain (BSG), a major byproduct of the beer industry, is rich in fibers, minerals, proteins and other valuable chemical compounds [2]. In the present study, spent malt grain (SMG), which contains similar compounds to BSG, was used and sourced from a non-alcoholic malt beverage production facility. The SSF has been interested due to its ability to grow micro-organisms on waste substrates under limited or non-free water conditions. This process can enhance the substrate by decreasing fiber and increasing protein and amino acid contents, making it appropriate as food or feed supplements for humans and animals. Solid-state fermentation of BSG using probiotic microorganisms includes the potential to produce products with beneficial effects on consumers [3].
Probiotics are defined as “live microorganisms that provide health benefits to the host once consumed in sufficient quantities” [4]. Use of probiotics not only plays a role in human health, but also enhances natural digestion and maintains health of animals. In addition, it is effective in preventing activity of pathogens, metabolism of nutrients, removal of toxic substances and energy balance [5]. Beneficial effects of probiotic microorganisms on the substrate are connected to their ability to tolerate conditions of the stomach and intestines in humans and animals, which completely depends on the type of microorganisms and their resistance to acid and bile. Probiotics such as Lactobacillus strains include proteins resistant to acid shock. Numerous microorganisms are susceptible to destruction by bile salts due to the presence of lipids and fatty acids in their cell membranes. However, many lactic acid bacteria (LABs) are resistant to bile [6, 7].
Fermentation of agricultural wastes using probiotic bacteria enhances antioxidant and antimicrobial activities of the fermented products and protects gut microbiota against oxidative stress [8]. Fermentation of cereal wastes with P. acidilactici inhibits growth of Aspergillus, Penicillium and Fusarium Sps. by producing antimicrobial compounds; thereby, preventing mycotoxin production. The resulting fermented products, rich in probiotic bacteria, can be used in human foods and animal feeds [9]. The complex lingo-cellulosic structure of agricultural wastes poses challenges for their use as substrates by microorganisms, especially bacteria. Therefore, various pretreatment methods are used to modify them. These methods, including physical, chemical, physicochemical, ionic liquid and biological approaches, have been studied for enhancing nutrient availability in BSG. Each method includes strengths and limitations. Physical pretreatments need high energy but may not fully degrade lignocellulose. Chemical methods improve nutrient accessibility but create inhibitory byprod-ucts that need neutralization. Ionic liquids offer effective breakdown but are expensive and challenging to recover. Physicochemical treatments are efficient but costly. Optimizing BSG physicochemical pretreatment processes can decrease energy consumption. This enhances efficiency of nutrient release and substrate degradation. Biological pretreatments with white and soft-rot fungi are environ-mental friendly and produce nutrient-rich substrates; how-ever, they are time-intensive. [10,11]. This method modifies lignocellulosic structure of the substrate by partially or completely removing lignin; thereby, enhancing microbial access to cellulosic parts of the raw materials. The primary focus includes breaking down lignin and hemi-cellulose; then, allowing it to digest cellulose effectively [12]. Use of fungi in fermentation process increases nutritional quality of the animal feeds prepared from BSG [13].
Carrying out and optimizing a pretreatment facilitate probiotic growth; by which, cellulosic and hemicellulosic structures of BSG are broken down. For example, A. oryzae is an appropriate option for the fungal pretreatment of BSG because of its lignocellulosic, proteolytic and amylolytic enzymes [14]. Treatment with A. oryzae results in produc-tion of fermentable sugars and increases in the substrate protein contents. When A. oryzae spores are directly used in BSG treatment, they can serve as a nutrient source in a probiotic propagation media [15]. In previous studies, SSF of BSG has majorly been carried out using fungi and a bacterial strain of Bacillus Sp. for purposes, including production of value-added fermented products, enzymes, lactic acid and proteins [13,16,17]. However, use of probiotic bacteria to enhance the nutritional value of this substrate has not been investigated. From the four probiotic bacterial strains assessed in the study (based on their resist-ance to acid and bile and antimicrobial, antitoxic and anti-oxidant activities), the best one was selected for investiga-ting growth on SMG and fungal pretreatment.
In this study, SSF of SMG was investigated for the first time using probiotic P. acidilactici and fungal pretreatment of A. oryzae PTCC 5163. Effects of this pretreatment on the growth and viability of the selected probiotic bacterium and modification of the chemical composition were investigated as well.
- Materials and Methods
2.1 Materials
Most chemicals and brain-heart infusion soft agar (BHI), Rogosa Sharpe broth (MRS broth), Rogosa, Sharpe agar (MRS agar), nutrient agar (NA) and nutrient broth (NB) were purchased from Merck, Darmstadt, Germany. Potato dextrose agar (PDA agar), ProMedia, bile salts and DPPH were purchased from Merck and Sigma-Aldrich, Darmstadt, Germany. Rice containing aflatoxin was provided by Department of Poultry Sciences, Faculty of Agriculture, Tarbiat Modares University, Tehran, Iran. The P. acidi-lactici is a commercial strain isolated from Bactocell© probiotic (Lallemand Animal Nutrition, France), L. plantarum MT. ZH393, L. fermentum MT. ZH893, lactobacilli isolated from Mazandaran cheese (isolated in Department of Food Science and Technology, Tarbiat Modares University), Escherichia coli Nissle 1917 isolated from Mutaflor prob-iotic, A. oryzae PTCC 5163, Salmonella serovar enteritidis RTCC 1621, Salmonella enterica serovar typhimurium RTCC 1679, Salmonella serovar infantis ATCC 51741, E. coli O1:K1, E. coli O2:K2 and E. coli O78:K80 (strain χ1378) were provided by the microbial collection of the faculty of veterinary medicine, university of Tehran, Tehran, Iran. The SMG was provided by Behnoosh, an alcohol-free beer manufacturer, Tehran, Iran.
2.2 Methods
2.2.1 Probiotic strains
Four probiotic strains were used in this study, including P. acidilactici, L. plantarum MT. ZH393, L. fermentum MT. ZH893 and E. coli Nissle 1917.
2.2.2 Preparation of microbial cultures
To activate the probiotic strains, except E. coli Nissle 1917, a small quantity of the lyophilized microbial powder was poured into 10 ml of sterile MRS broth. Mixture was incubated at 37 ˚С for 24 h under anaerobic conditions. Nutrient broth was used for the growth of E. coli Nissle 1917. Overnight cultures of probiotic strains were centrifug-ed at 6000× g for 10 min at 4 °C (Model 3-30KS, Sigma, Germany). Cells separated from the culture media were washed twice with sterile Ringer's solution and then suspended in Ringer's solution. Suspensions were prepared to 0.5 McFarland standard (≃1.5×108 CFU.ml-1) and inocul-ated to the substrate at an inoculum culture volume of 20% of the total substrate (v.w-1). Methods of preparing microbial suspension were similar for all the experiments of the study needing microbial suspension. Pathogenic indicator bacteria were activated in Mueller-Hinton broth for 24-48 h at 37 °C. To prepare stock culture of A. oryzae PTCC 5163 spores, the microorganism was cultivated on the surface of PDA. Spores were formed and collected after 10-12 d of incubation at 25 °C. Spores were passed through cotton filters to separate mycelia and then diluted with a phosphate buffer saline (PBS) solution (pH 7). The number of spores was adjusted to 1.6×107 spores.ml-1 using a hemacytometer slide (MarienFeld, Germany). Then, sterile glycerol 10-13% )vol.) was added to the mixture. This was divided into small vials and stored at -20 °C until use.
2.2.3 Antimicrobial activity
Agar spot test was used to assess the ability of strains to inhibit the growth of pathogenic microbes (Salmonella serovar enteritidis RTCC 1621, Salmonella serovar typhim-urium RTCC 1679, Salmonella serovar infantis ATCC 51741, E. coli O1:K1, E. coli O2:K2 and E. coli O78:K80 (strain χ1378). Two microliters of a 24-h culture of L. plantarum MT. ZH393, L. fermentum MT ZH893, and P. acidilactici were spotted on the surface of MRS agar and E. coli Nissle 1917 strain was spotted on nutrient agar. All plates were incubated anaerobically at 37 °C for 24 h. Then, 10 ml of BHI soft agar (0.7%) were mixed with 100 µl of the activated pathogenic microorganisms and spread on the surface of MRS agar. Media were incubated at 37 °C for 24 h [18]. Presence of a clear zone or halo less than 1 mm was classified as +, while a halo of 2–5 mm was categorized as ++, a halo greater than 5 mm or complete inhibition of growth was categorized as +++ and absence of any halo formation was categorized as –.
2.2.4 Resistance to acidic conditions
Briefly, L. plantarum MT, ZH393, L. fermentum MT. ZH893 and P. acidilactici in MRS broth and E. coli Nissle 1917 in nutrient broth were incubated at 37 °C for 48 h. After assessing population of the microorganisms, 1 ml of each medium was inoculated into 9 ml of PBS (pH 7.4) with pH adjusted to 2.5 using HCl. After 2 h of incubation at 37 °C, number of the micro-organisms was re-enumerated. Results were compared to assess effects of acidic environ-ment on decreasing population of probiotic strains [18, 19].
2.2.5 Bile resistance
Bile resistance of the microorganisms was assessed using a method by Sharifi Yazdi et al; in which, 100 µl of the microbial suspension (prepared as described in Section 2.2.2) were added to the culture media with bile salts (Model B8756, Sigma-Aldrich, Germany) and without bile salts, respectively. Then, light absorption of the samples was measured using a spectrophotometer (Model Cary 60 UV-Vis, Agilent Technologies, USA) at 600–650 nm before and after 8 h of incubation [20]. The inhibition coefficient (Cinh) was calculated using the Eq. 1.
Cinh = (T8 - T0) control - (T8 - T0) treatment) / (T8 - T0) control Eq. 1
Where, T0 and T8 included reading at 0 and 8 h, respectively.
2.2.6 Antioxidant activity
One milliliter of each microbial suspension was added to 1 ml of a newly prepared DPPH solution (2,2-diphenyl-1-picrylhydrazyl). Then, mixture was agitated strenuously and set for 30 min at ambient temperature in dark. After re-centrifugation (8000× g, 10 min, 4 °C), decreases in absorb-ance were measured at 517 nm [8,21]. The scavenging activity was calculated using Eq. 2.
Scavenging activity (%) = 1 - (Asample - Ablank /Acontrol) × 100 Eq. 2
Where, sample included cells, DPPH, and methanol, blank included cells and methanol, and control included Ringer’s solution and DPPH.
2.2.7 Antitoxic characteristics
In this study, the toxin included aflatoxin. The best food method was used to extract aflatoxins from contaminated rice based on the AOAC official method [22]. Generally, 1 g of the toxin-containing sample was mixed with 20 ml of 55% (vol.) aqueous methanol and agitated for 30 min. Then, 10 ml of hexane were added to the mixture and agitated for 15 min. Then, mixture was centrifuged at 2000× g for 5 min and the supernatant was discarded. Then, 20 ml of chloroform were added to the mixture and agitated gently for 15 min. The lower phase was filtered using filter paper coated with anhydrous sodium sulfate (1 g in the bottom of the funnel) and the toxin dissolved in chloroform was concentrated by evaporation of chloroform. The extracted toxin was dissolved in 5 ml of 55% (vol.) aqueous methanol and added drop by drop to 20 ml of phosphate buffer solution saline (pH 7) until the absorbance of 360 nm reached nearly 0.1 (initial absorption). A microbial suspension was prepared from each strain overnight culture with a concentration of 8 McFarland (2.4 × 109 CFU.ml-1). Then, 1 ml of the microbial suspension was poured into a 2-ml microtube and centrifuged at 10000× g for 10 min (Heraeus Biofuge Pico, Thermo Fisher Scientific, USA). Supernatant of the microbial pellets was discarded and the pellets were washed twice with PBS solution (pH 7). One milliliter of the prepared toxin was added to the pellets in the bottom of the microtubes. Toxin and pellets were incubated for 4-5 min and thoroughly mixed. They were incubated for 1 h, absorbance was read at 360 nm and the absorption decrease percentage was calculated through Eq. 3.
Eq. 3
2.2.8 Spent malt grain preparation
Wet SMG from a malt beverage factory was dried at 50 °C for 24 h using the oven (Model NST-F300-51605, Noor Sanat Ferdows, Iran). After reaching a constant weight and grinding, this was passed through a 1-mm sieve and stored at 4 °C for further use.
2.2.9 Preparation of spent malt grains for solid-state fermentation
Briefly, 10 g of SMG was poured into a 250-ml Erlen-meyer flask. The moisture was adjusted to 70% based on dry matter and pH was adjusted to 5.4 using calcium carbonate solution (2.5% w.v-1), which was an appropriate pH for the growth of A. oryzae PTCC 5163. Then, this was sterilized at 121 °C for 15 min using autoclave (Model 121 A, Iran Tolid, Iran).
2.2.10 Fungal pretreatment
Briefly, 2 ml of A. oryzae PTCC 5163 spores (1.6 × 107 spores.ml-1) were inoculated into sterile SMG, mixed well and incubated at 30 °C for 24 h. Sterile calcium carbonate solution (2.5% w.v-1) was used to adjust the pH to 6.15–6.2. It is noteworthy that all steps were carried out under completely sterile conditions. Then, 2 ml of a P. acidilactici suspension (prepared as described in the Preparation of Microbial Cultures section) adjusted to 0.5 McFarland were inoculated into the sample; in which, A. oryzae PTCC 5163 was grown for 24 h. Flasks were sealed with parafilm and incubated at 37 °C for 24 h. To assess effects of fungal pretreatment, a series of experiments were carried out that involved inoculating the probiotic bacteria without prior fungal treatments.
2.2.11 Composition analysis of spent malt grains
Crude protein content was assessed using Kjeldahl method and the nitrogen content was multiplied by 6.25. Moreover, Soxhlet extraction was used to assess the lipid content (Model PSU-500S, Pecofood, Iran). To assess the ash content, samples were burned in electric furnaces at 650 °C for 4 h (Model SIC 37, Ecotec, Iran). Moreover, the moisture content was estimated using weight differences at 105 °C for 3.5 h using oven (PAAT ARIA, Iran). All the assays were carried out according to AOAC [23]. Acid detergent fiber (ADF), neutral detergent fiber (NDF) and acid detergent lignin (ADL) were assessed using sodium sulfite and necessary solutions, based on the Van Soest method [24]. Hemicellulose was investigated from the differences between NDF and ADF and cellulose was investigated from the differences between ADF and ADL [17].
2.2.12 Statistical analysis of data
To assess significant differences between the means, one-way analysis of variance was carried out using Duncan's multiple range test. All experiments were carried out in triplicate, with a significance level set at p < 0.05. Additionally, figures were created using Excel 2016 software (Microsoft, USA).
- Results and Discussion
3.1 Antimicrobial activity
Among the probiotic strains, P. acidilactici completely prevented growth of the pathogenic strains. Moreover, L. fermentum MT. ZH893 and L. plantarum MT. ZH393 included good antimicrobial characteristics, compared to E. coli Nissle 1917 which included a weaker antimicrobial activity than other strains (Table 1, Figure 1). Metabolites such as bacteriocins produced by P. acidilactici might inhibit other LAB strains. Moreover, lactic and acetic acids from the fermentation process led to decreases in pH below 4.5; thus, preventing growth of pathogenic bacteria [9]. In a study by Tavakoli et al. strains of L. plantarum and L. fermentum showed desirable antimicrobial activities due to their metabolites e.g. organic acids, bacteriocins, and hydrogen peroxide [18]. In another study, it was indicated that metabolites of P. acidilactici such as bacteriocins demonstrated antimicrobial activities against three indicator bacteria of L. monocytogenes, E. coli and S. aureus [25].
3.2 Resistance to acidic conditions
From the four strains, only E. coli Nissle 1917 showed significantly lower resistance to acidic conditions, compared to those the other strains did. The other probiotic strains demonstrated excellent acid resistance with no significant differences. Survival rates in acidic conditions for L. fermentum MT. ZH893 and P. acidilactici were assessed as 96 and 95.7%, respectively (Figure 2). Regarding resistance to acidic conditions, findings of this study were similar to those of a study; in which, P. acidilactici showed appropriate resistance to stomach acid conditions. This was possibly because of its ability to decrease activity of the H+-ATPase enzyme, which regulated internal and external pH and led to the resistance of microorganisms to acidic conditions [26]. In a study by Tavakoli et al., Lactobacillus strains showed resistance to acidic conditions and survival of the strains in acidic conditions varied 68.8–94.9% [18]. Several theories are available on the resistance of Gram-positive bacteria such as LAB to acidic conditions, including stability of their mRNA as well as systems responsible for modifying composition of the cell membrane, extruding protons, protecting macromolecules, changing metabolic pathways and producing alkali. Furthermore, proton pumps heavily rely on the F1F0-ATPase to maintain internal pH [27].
3.3 Bile resistance
Results of the present study demonstrated that all strains included good resistance to bile salts, however, inhibitory coefficient of P. acidilactici was the lowest, thus its resistance to bile was the highest (Figure 3). Resistance to bile in the probiotic strains might be explained by its special bilayer structure that could tolerate unfavorable alkaline conditions [26]. Probiotic strains resistant to bile salts can survive in the digestive system, form colonies and subsequently provide health benefits for humans and animals. Probiotic survival against bile balances the gut microorganisms and enhances overall health and performance [28]. Similarly, another report revealed that six probiotic isolates, belonging to P. acidilactici, included good bile resistance [20].
3.4 Antioxidant activity
Results indicated that P. acidilactici and L. fermentum MT. ZH893 included the highest scavenging rates (nearly 65%), compared to L. plantarum MT. ZH393 with the lowest rate (49%) (Figure 4). Regarding good antioxidant activity of the probiotic strains especially P. acidilactici, membrane-bound enzymes might be responsible for the antioxidant activity. One of these enzymes is dipeptidyl peptidase-III (DPP-III), which decreases oxidative stress and its associated diseases [29]. Li et al. reported that the highest quantity of scavenging activity for L. plantarum strains, isolated from Chinese fermented food, was roughly 50%; similar to the findings of the present study. Based on their study, proteins or polysaccharides on the cell surface contributed to the antioxidant activity of this strain since removing these compounds decreased capacity to remove DPPH free radicals [21]. In another study, a wild L. plantarum strain isolated from fermented cabbages significantly improved scavenging activities of superoxide anion, DPPH and hydroxyl radicals. Naturally, these radicals contribute to lipid oxidation, as they are precursors of singlet oxygen and hydroxyl radicals [30].
3.5 Antitoxic characteristics
Results of the present study demonstrated that all probiotics could bind to aflatoxins, although two strains of P. acidilactici and, L. fermentum MT. ZH893 showed greater absorption rates. Moreover, no significant difference was seen at 5% level with L. plantarum MT. ZH393 (Figure 5).
Studies have shown strain-dependent relationships between toxin binding and LAB. Furthermore, it was demonstrated that polysaccharides and peptidoglycans of the bacterial cell walls possibly contributed to aflatoxin binding [31]. Since probiotic bacteria attach to aflatoxins through physical adhesion rather than covalent bonding, non-viable bacterial cells can bind to them as well. In other words, probiotic microorganisms (bacteria and fungi) and their enzymatic metabolites detoxify aflatoxin molecules by cleaving their difuran rings [32]. Similarly, it is reported that L. plantarum isolated from a type of fermented cheeses demonstrated a strong ability to bind to aflatoxin B1 when the microorganism was alive and after it was killed by heat [33].
3.6 Fungal pretreatment
After selecting P. acidilactici based on its superior probiotic characteristics, effects of fungal pretreatment with A. oryzae PTCC 5163 on its growth and survival were studied. Results of the present study indicated that fungal pretreatment at various pH levels included significant differences in the number of probiotics, compared to the control condition (without fungal pretreatment). Adjusted pH values were assessed based on the pH range appropriate for the growth of fungi and bacteria. At all pH values, pretreatment by A. oryzae PTCC 5163 significantly increased the number of P. acidilactici after 48 h of fermentation (from 9.4 to 9.76 log CFU. G-1) (Figure 6). The BSG is full of lignocellulosic materials that can be destroyed by filamentous fungi, whose hyphae can easily penetrate the inter-particle space. Therefore, using fungi for pretreatment is an appropriate alternative to chemical and physical methods because in addition to be cost-effectiveness, they can easily be used in SSF [13].
The P. acidilactici can convert cereal byproducts into products that can be used in human and animal foods [9]. Enhancing the microbial growth and survival yields further favorable outcomes in the substrate. Pretreatment of lignocellulosic materials enhances accessibility of the substrate for probiotic bacteria because of the hydrolysis of hemicellulose, cellulose and lignin. Fungal pretreatment by Aspergillus strains with amylolytic and cellulolytic enzymes is an environmental friendly method that increases access of probiotic bacteria to materials needed for their growth [34, 35]. A study showed that use of A. oryzae and S. cerevisiae could produce bioethanol from BSG. Technically, A. oryzae breaks down cellulose and hemicellulose, creating sugars for ethanol production by S. cerevisiae. This pretreatment provides necessary enzymes without excessive sugar breakdown or high levels of furan-based inhibitors [36].
3.7 Assessing spent malt grain compounds
Fermentation of SMG with P. acidilactici following fungal pretreatment with A. oryzae PTCC 5163 led to significant increases in protein contents with 32.4% increases (Table 2). Increases in protein during fermentation could include various causes. One possible reason included the accumulation of fungal and bacterial biomasses in SMG substrate after fermentation [17]. These results were similar to results reported by Bekatorou et al., who reported 20–36% increases in the protein contents of BSG, when treated by Aspergillus strains [15]. In the fermentation of brewer's dried grains and spent sorghum using microorganisms, more than 30% increases in protein contents were reported [38]. The lipid contents of SMG did not change significantly after fermentation (Table 2). This finding was similar to those of other studies, where fermentation with LAB did not lead to significant changes in lipid contents [8].
A significant result of the fermentation process included decrease in fiber contents. The NDF and ADF decreased by 17.7 and 8.3%, respectively. Additionally, cellulose and hemicellulose contents decreased by 27.35 and 8.5%, respectively (Table 2). This decrease in fiber contents could be attributed to activity of the microorganisms and their metabolites, breaking down complex carbohydrates such as cellulose and hemicellulose. These findings are supported by previous studies; in which, fermentation with LAB and their metabolites such as organic acids and bacteriocins led to decreases in fiber contents. Similarly, SSF of BSG with fungi have been shown to decrease fiber and cellulose contents [37]. Fermentation process resulted in a significant increase of 59.6% in ash contents (Table 2). Increase in ash contents might reflect accumulation of minerals and other inorganic components in fermented SMG or was likely due to metabolic activity of the microorganisms and their metabolites during fermentation [17]. Similar studies reported significant increases in ash contents following fermentation of agricultural wastes and BSG by bacteria and fungi [38].
- Conclusion
The study reported that P. acidilactici included superior antimicrobial characteristics and bile resistance, compared to those the L. plantarum MT. ZH393, L. fermentum MT. ZH893 and E.coli Nissle 1917 did. Its antitoxic and anti-acidic characteristics did not show significant differences at 5% level. The SSF resulted in lower bacterial growth than that fungi did, but pretreatment with fungi significantly boosted development of the probiotic bacterial strain. Combination of fungal pretreatment and SSF led to changes in the culture media composition, including decreases in fibers (17.7%) and increases in proteins (32.4%) and ashes (59.6%). Decreasing fibers enhances the product's bioavailability; thereby, improving access to other essential nutrients. Significant increases in protein contents transform the products into potential protein sources. Moreover, increased ash contents signified higher mineral concen-trations within the fermented products. This study innovatively used solid-state fermentation of SMG with P. acidilactici, using A. oryzae fungal pretreatment to enhance bacterial growth, marking a novel approach in producing high-value fermented products. The fermented product includes the potential for use in livestock and poultry feeds or as a dietary supplement in associated industries.
- Acknowledgements
The financial support from the Research Council of Tarbiat Modares University (IG-39804) is gratefully acknowledged. The authors express their deepest gratitude to the Faculty of Technical and Natural Resources of Tuyserkan College, Bu-Ali Sina University, for providing facilities for some experiments.
- Conflict of Interest
All authors declare no conflict of interest.
- Authors Contributions
“Conceptualization, Hamidi-Esfahani Z; methodology, Hamidi-Esfahani Z and Karimi-Torshizi MA; validation, Hamidi-Esfahani Z, Shokouhi M and Karimi-Torshizi MA; investigation, Shokouhi M; writing-original draft preparation, Shokouhi M; writing-review and editing, Hamidi-Esfahani Z; supervision, Hamidi-Esfahani Z and Karimi-Torshizi MA”
- Antimicrobial activity
- Antitoxic characteristics
- Biological pretreatment
- Brewer’s spent grain (BSG)
- Probiotic bacteria
- Survival rate
How to Cite
References
Sadh PK, Duhan S, Duhan JS. Agro-industrial wastes and their utilization using solid state fermentation: A review. Bioresour Bioprocess. 2018; 5(1): 1-5. https://doi.org/10.1186/s40643-017-0187-z
Lisci S, Tronci S, Grosso M, Karring H, Hajrizaj R, Errico M. Brewer's spent grain: Its value as renewable biomass and its possible applications. Chem Eng Trans. 2022;92:259-264. https://doi.org/10.3303/CET2292044
Lock TJ, Mah SH, Lai ZW. Versatile applications of brewer’s spent grain: Solid-state fermentation and nutritional added value. Appl Biochem Biotechnol. 2024; 196(8): 5508-5532. https://doi.org/10.1007/s12010-023-04769-3
Hotel ACP, Cordoba A. Health and nutritional properties of probiotics in food including powder milk with live lactic acid bacteria. Prevent. 2001; 5(1): 1-10.
Bubnov RV, Babenko LP, Lazarenko LM, Mokrozub VV, Spivak MY. Specific properties of probiotic strains: relevance and benefits for the host. EPMA J. 2018; 9(2): 205-223. https://doi.org/10.1007/s13167-018-0132-z
Jin LZ, Ho YW, Abdullah N, Jalaludin S. Acid and bile tolerance of Lactobacillus isolated from chicken intestine. Lett Appl Microbiol. 1998; 27(3): 183-185. https://doi.org/10.1046/j.1472-765x.1998.00405.x
Song M, Yun B, Moon JH, Park D-J, Lim K, Oh S. Characterization of selected Lactobacillus strains for use as probiotics. Korean J Food Sci Anim Resour. 2015; 35: 551-556. https://doi.org/10.5851/kosfa.2015.35.4.551
Mladenović D, Pejin J, Kocić-Tanackov S, Djukić-Vuković A, Mojović L. Enhanced lactic acid production by adaptive evolution of Lactobacillus paracasei on agro-industrial substrate. Appl Biochem Biotechnol. 2019; 187: 753-769. https://doi.org/10.1007/s12010-018-2852-x
Bartkiene E, Bartkevics V, Krungleviciute V, Juodeikiene G, Zadeike D, Baliukoniene V, Bakutis B, Zelvyte R, Santini A, Cizeikiene D. Application of hydrolases and probiotic Pediococcus acidilactici BaltBio01 strain for cereal byprod-ucts conversion to bioproduct for food/feed. Int J Food Sci Nutr. 2018; 69(2): 165-175.
Mitri S, Salameh SJ, Khelfa A, Leonard E, Maroun RG, Louka N, Koubaa M. Valorization of brewers’ spent grains: Pretreatments and fermentation, a review. Ferment. 2022; 8(2): 50. https://doi.org/10.3390/fermentation8020050
Castilla-Archilla J, Cermeño M, Tuohy MG, FitzGerald RJ, Lens PN. Brewers’ spent grain pretreatment optimisation to enhance enzymatic hydrolysis of whole slurry and resuspended pellet. Front Chem Eng. 2023; 12(5): 1272988. https://doi.org/10.3389/fceng.2023.1272988
Mishra S, Singh PK, Dash S, Pattnaik R. Microbial pretreatment of lignocellulosic biomass for enhanced biomethanation and waste management. 3 Biotech. 2018; 8: 1-12. https://doi.org/10.1007/s13205-018-1480-z
Marcus A, Fox G. Fungal biovalorization of a brewing industry byproduct, brewer’s spent grain: A review. Foods. 2021; 10(9): 2159.https://doi.org/10.3390/foods10092159
Wilkinson S, Smart KA, Cook DJ. Optimisation of alkaline reagent based chemical pre-treatment of Brewers spent grains for bioethanol production. Ind Crops Prod. 2014; 62: 219-227. https://doi.org/10.1016/j.indcrop.2014.08.036
Bekatorou A, Bountas Y, Banat IM, Kanellakl M. Upgrading brewer's spent grains by treatment with Aspergillus species. Chem Ind Chem Eng Q. 2007; 13(2): 72-78.https://doi.org/10.2298/CICEQ0702072B
Tan, Y. X., Mok, W. K., Lee, J., Kim, J., Chen, W. N. Solid state fermentation of Brewers’ spent grains for improved nutritional profile using Bacillus subtilis WX-17. Fermentation. (2019); 5(3): 52. https://doi.org/10.3390/fermentation5030052
Terrasan CRF, Carmona EC. Solid-state fermentation of brewer's spent grain for xylanolytic enzymes production by Penicillium janczewskii and analyses of the fermented substrate. Biosci J. 2015; 31(6): 1826-1836. https://doi.org/10.14393/BJ-v31n6a2015-30044
Tavakoli M, Hamidi-Esfahani Z, Hejazi MA, Azizi MH, Abbasi S. Characterization of probiotic abilities of Lactobacilli isolated from Iranian Koozeh traditional cheese. Pol J Food Nutr Sci. 2017; 67(1): 41-48. https://doi.org/10.1515/pjfns-2016-0003
Guo Z, Wang J, Yan L, Chen W, Liu X-m, Zhang H-p. In vitro comparison of probiotic properties of Lactobacillus casei Zhang, a potential new probiotic, with selected probiotic strains. LWT-Food Sci Technol. 2009; 42(10): 1640-1646. https://doi.org/10.1016/j.lwt.2009.05.025
Sharifi Yazdi MK, Davoodabadi A, Khesht Zarin HR, Tajabadi Ebrahimi M, Soltan Dallal MM. Characterisation and probiotic potential of lactic acid bacteria isolated from Iranian traditional yogurts. Ital J Anim Sci. 2017; 16(2): 185-188. https://doi.org/10.1080/1828051X.2016.1222888
Li S, Zhao Y, Zhang L, Zhang X, Huang L, Li D, Niu C, Yang Z, Wang Q. Antioxidant activity of Lactobacillus plantarum strains isolated from traditional Chinese fermented foods. Food Chem. 2012; 135(3): 1914-1919. https://doi.org/10.1016/j.foodchem.2012.06.048
Official Methods of Analysis of AOAC International, 17th ed., Method 975.36. Aflatoxins in Food and Feed. AOAC International, Gaithersburg, MD, 2000.
Lee J, Durst R, Wrolstad R. AOAC official method 2005.02: total monomeric anthocyanin pigment content of fruit juices, beverages, natural colorants and wines by the pH differential method. Official methods of analysis of AOAC International. 2005;2.
Van Soest Pv, Robertson JB, Lewis BA. Methods for dietary fiber, neutral detergent fiber and nonstarch polysaccharides in relation to animal nutrition. J Dairy Sci. 1991; 74(10): 3583-3597. https://doi.org/10.3168/jds.S0022-0302(91)78551-2
Sidek NLM, Halim M, Tan JS, Abbasiliasi S, Mustafa S, Ariff AB. Stability of bacteriocin-like inhibitory substance (BLIS) produced by Pediococcus acidilactici kp10 at different extreme conditions. BioMed Res Int. 2018; 2018. https://doi.org/10.1155/2018/5973484
Olajugbagbe TE, Elugbadebo OE, Omafuvbe BO. Probiotic potentials of Pediococuss acidilactici isolated from wara; A Nigerian unripened soft cheese. Heliyon. 2020; 6(9). https://doi.org/10.1016/j.heliyon.2020.e04889
Matsumoto M, Ohishi H, Benno Y. H+-ATPase activity in Bifidobacterium with special reference to acid tolerance. Int J Food Microbiol. 2004; 93(1): 109-113. https://doi.org/10.1016/j.ijfoodmicro.2003.10.009
Shokryazdan P, Faseleh Jahromi M, Liang JB, Ho YW. Probiotics: from isolation to application. J Am Coll Nutr. 2017; 36(8): 666-676. https://doi.org/10.1080/07315724.2017.1337529
Attri P, Jodha D, Gandhi D, Chanalia P, Dhanda S. In vitro evaluation of Pediococcus acidilactici NCDC 252 for its probiotic attributes. Int J Dairy Technol. 2015; 68(4): 533-542. https://doi.org/10.1111/1471-0307.12194
Gao D, Gao Z, Zhu G. Antioxidant effects of Lactobacillus plantarum via activation of transcription factor Nrf2. Food Funct. 2013; 4(6): 982-989. https://doi.org/10.1039/c3fo30316k
Bagherzadeh Kasmani F, Torshizi K, Allameh AA, Shariatmadari F. Aflatoxin detoxification potential of lactic acid bacteria isolated from iranian poultry. Iran J Vet Res. 2012; 13(2): 152-155. https://sid.ir/paper/592937/en
Afshar P, Shokrzadeh M, Raeisi SN, Ghorbani-HasanSaraei A, Nasiraii LR. Aflatoxins biodetoxification strategies based on probiotic bacteria. Toxicon. 2020; 178: 50-58. https://doi.org/10.1016/j.toxicon.2020.02.007
Huang L, Duan C, Zhao Y, Gao L, Niu C, Xu J, Li S. Reduction of aflatoxin B1 toxicity by Lactobacillus plantarum C88: a potential probiotic strain isolated from Chinese traditional fermented food “tofu”. PloS ONE. 2017; 12(1): e0170109. https://doi.org/10.1371/journal.pone.0170109
Lee K, Lee SK, Lee BD. Aspergillus oryzae as probiotic in poultry-A review. Int J Poult Sci. 2006; 5(1): 1-3. https://doi.org/10.3923/ijps.2006.1.3
Hui L, Wan C, Hai-Tao D, Xue-Jiao C, Qi-Fa Z, Yu-Hua Z. Direct microbial conversion of wheat straw into lipid by a cellulolytic fungus of Aspergillus oryzae A-4 in solid-state fermentation. Bioresour Technol. 2010; 101(19): 7556-7562. https://doi.org/10.1016/j.biortech.2010.04.027
Wilkinson S, Smart KA, James S, Cook DJ. Bioethanol production from brewers spent grains using a fungal consolidated bioprocessing (CBP) approach. Bioenergy Research. 2017; 10: 146-157. https://doi.org/10.1007/s12155-016-9782-7.
Estevão-Rodrigues T, Fernandes H, Moutinho S, Filipe D, Fontinha F, Magalhães R, Couto A, Ferreira M, Gamboa M, Castro C, Belo I. Effect of solid-state fermentation of brewer's spent grain on digestibility and digestive function of European seabass (Dicentrarchus labrax) juveniles. Anim Feed Sci Technol. 2024; 315: 116018. https://doi.org/10.1016/j.anifeedsci.2024.116018
Abasiekong S. Effects of fermentation on crude protein content of brewers dried grains and spent sorghum grains. Bioresour Technol. 1991; 35(1): 99-102. https://doi.org/10.1016/0960-8524(91)90088-2
- Abstract Viewed: 429 times
- pdf Downloaded: 146 times