Assessment of Probiotic Characteristics of Lactiplantibacillus plantarum SU-KC1a Isolated from Human Breast Milk in Indonesia
Applied Food Biotechnology ,
Vol. 11 Núm. 1 (2024),
18 noviembre 2023
,
Página e31
https://doi.org/10.22037/afb.v11i1.45707
Resumen
Background and Objective: Lactic acid bacteria such as Lactiplantibacillus spp. and Bifidobacterium spp. are addressed for their beneficial characteristics. This study aimed to assess probiotic attributes of Lactiplantibacillus plantarum SU-KC1a isolated from human breast milk in Indonesia.
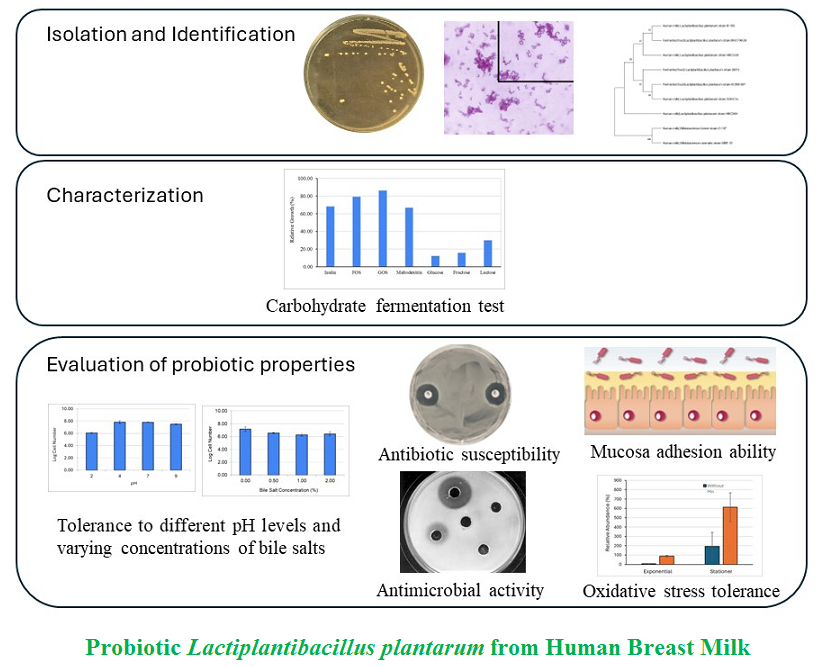
Material and Methods: Lactiplantibacillus plantarum SU-KC1a was anaerobically isolated from human breast milk and then morphologically and biochemically characterized. Probiotic characteristics, including tolerance to various pH levels and bile salt concentrations, antibiotic susceptibility, antimicrobial activity, mucosal adhesion ability and oxidative stress tolerance, were assessed.
Results and Conclusion: Lactiplantibacillus plantarum SU-KC1a exhibited rod-shaped, Gram-positive staining, negative catalase activity, and lacked acid-fastness and motility. Under stress conditions, this strain adopted a coccoid-like shape to protect its cell membrane and internal structures from external environments. Significantly, it demonstrated resistance to mupirocin, distinguishing it from other lactobacilli. The strain showed enhanced growth in the presence of prebiotics, particularly galactooligosaccharides. Additionally, it showed probiotic characteristics such as robust tolerance to pH variations and bile salts, potent antimicrobial activity against pathogens, strong adherence to mucosal surfaces and resistance to oxidative stress. These findings highlight Lactiplantibacillus plantarum SU-KC1a as a promising probiotic candidate appropriate for incorporation into functional foods.
Conflict of interest: The authors declare no conflict of interest.
- Introduction
Breast milk is renowned for its rich composition of essential nutrients, bioactive compounds and antibodies critical for infant growth and immune development. It contains a spectrum of elements, including carbohydrates, proteins, antibodies, vitamins, minerals and amino acids (notably L-arginine), making it a perfect natural nourishment for infants [1]. Studies have consistently highlighted advantages of breastfeeding, showing a lower susceptibility to infections and gastrointestinal disorders in breast-fed neonates, partly due to the presence of probiotics in breast milk [1, 2]. From these probiotics, lactic acid bacteria (LAB) such as Lactiplantibacillus spp. and Bifido-bacterium spp. are particularly prominent for their beneficial effects on the infants’ immune system. Naturally, LAB play their significant roles in promoting human health by stimulating proliferation and differentiation of the host intestinal cells, regulating acidity in the gastrointestinal tract (GIT) and competing with harmful pathogens [3].
Breast milk contains prebiotics known as human milk oligosaccharides to increase growth and viability of LAB. Importance of prebiotics in breast milk is signified in industrial practices, where prebiotics are intentionally introduced to functional foods to support LAB growth and viability. This integration of prebiotics and probiotics in food production serves to nourish beneficial bacteria, enhancing their survival and functionality.
Oligosaccharides widely used as prebiotics in food industries include fructooligosaccharides (FOS) and galactooligosaccharides (GOS), lactose, isomaltooligo-saccharides, xylooligosaccharides, maltodextrin and inulin [4]. In addition to its prebiotic characteristics, inulin is used to improve the quality of milk fermented by Lactobacillus acidophilus and Bifidobacterium lactis [5]. Although addition of prebiotics can stimulate growth of LAB, those used in food industries often facing challenging environ-ments, resulting in decreased viabilities during processing and storage. As many LAB are facultative anaerobes or microaerophiles, they are highly susceptible to oxidative stress. Zhai et al. [6] reported that the growth of L. plantarum strain CAUH2 decreased when exposed to hydrogen peroxide. Another study by Dosan et al. [7] revealed that B. animalis subsp. lactis BR2-5 and B. breve BS2-PB3 cells were significantly depleted when exposed to oxygen, with no viable cells detected under aerobic conditions.
Categorized as probiotics, microorganisms must with-stand stressful conditions, exhibit survival capability within GIT, show effective adherence to intestinal epithelia and be safe for animals and humans [8]. Adherence of probiotic bacteria to the intestinal lining includes particular significance due to its role in the bacterial survival within the dynamic digestive tract environment. Colonization, a critical aspect of this survival, is facilitated through bacterial adhesion to the intestinal wall [9]. Additionally, LAB, specifically L. plantarum strain WLPL04 isolated from human breast milk (HBM), has been reported for its capacity to prevent adhesion of pathogenic bacteria via competitive mechanisms. These inhibitory effects have been demonstrated in vitro against pathogens such as Escherichia coli O157:H7, Salmonella Typhimurium ATCC13311 and Staphylococcus aureus CMCC26003 [10].
Despite extensive global studies on probiotic potential of LAB, a significant gap remains in understanding their isolation and characterization from HBM samples across diverse geographical regions. Studies focusing on LAB from HBM in various regions of the world, including Indonesia, are significantly limited. This gap is critical as regional variations in microbial composition can affect beneficial characteristics of LAB, especially in promoting maternal-infant health. Addressing this gap is essential for gaining a comprehensive understanding of LAB diversity and their potential uses in nutrition and health worldwide. Therefore, this study aimed to isolate and characterize LAB strains from HBM samples in Indonesia. Through compre-hensive biochemical profiling, molecular identification, assessment of growth capabilities on prebiotic substrates and assessment of the probiotic attributes of the identified strains, this study provided an overview of potential probiotic stains from HBM that could improve public health globally.
- Materials and Methods
2.1. Isolation and identification
2.1.1 Subjects and ethical issue
All procedures were approved by the Ethical Committee of Rumah Sakit Anak dan Bunda Harapan Kita, Jakarta, Indonesia (922/2020 on 03/31/2020) and all the volunteers signed written informed consents. Overall, two healthy women with no underlying conditions or complications participated in this study. They provided breast milk samples 1–2 d after their caesarean deliveries.
2.1.2 Sample collection
Sampling method was adapted from Eglash and Simon [11] with some modifications. All the equipment for sampling were sterilized for 10 min in boiling water and dried at room temperature (RT) before use. Nipples and mammary areola were cleaned with sterile water and cotton soaked in chlorhexidine (0.1%). The first drops were discarded, then the milk samples were collected using electric breast pump (Real Bubee, UK) and then transferred to sterile breast-milk storage bags. All samples were stored in a cool box with ice packs during transport to the laboratory.
2.1.3 Media
Two different media were used in this study, including de Man, Rogosa and Sharpe (MRS) (Liofilchem, Italy) and trypticase phytone yeast extract (TPY). One liter of TPY broth was prepared by mixing 17.6 g of tryptone soya broth (TSB) (Oxoid, UK), 5 g of peptone (Merck, USA), 5 g of glucose (Merck, USA), 2.5 g of yeast extract (Merck, USA), 1 ml of Tween-80 (Merck, USA) and 2 g of dipotassium phosphate (Merck, USA) in distilled water (DW). The two media were modified by the addition of L-cysteine (2.5 g/l) (Nowfood, USA) and mupirocin (0.05 mg/ml) (Genero, UK). Modified MRS and TPY were referred to as mMRS and mTPY, respectively. To prepare solid media, 1.5% bacteriological agar was added to the media.
2.1.4 Isolation of lactic acid bacteria
Milk samples were diluted using sterile peptone water (0.1%) (Merck, Germany) and spread-plated (50 µl) on media. To support growth of LAB at the isolation stage, mMRS agar supplemented with 30% pasteurized cow milk (Greenfields, Indonesia) was used. All plates were incubated at 37 oC for 96 h in an anaerobic jar (Oxoid, UK) with anaerobic atmosphere generator sachets (Oxoid, UK). After incubation, a colony was selected and purified using four-way streak method with the purified isolate referred to as SU-KC1a.
2.1.5 Molecular identification and phylogenetic analysis
For molecular identification, isolate was processed by PT Genetika Science Indonesia for Species Barcoding (DNA extraction, 16S rRNA gene amplification and molecular identification based on 16S rRNA sequence). The PCR reactions were prepared using universal primers 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-GGTTACCTTGTTACGACTT-3′). The 16S rRNA sequence was analyzed using Basic Local Alignment Search Tool (BLASTN 2.12.0+). Then, a phylogenetic tree was constructed using MEGA 11. MUSCLE alignment was carried out and a maximum likelihood phylogenetic tree was constructed. Bootstrap values were set at 1000 replicates. Sequences of other microorganisms used in the phylogenetic tree were retrieved from GenBank.
2.2 Characterization
2.2.1 Morphological study and biochemical activity test
The isolate was characterized based on morphology (Gram and acid-fast staining), motility and biochemical activity (catalase test). To assess effects of oxygen on cell morphology, the isolate was cultured on mMRS agar plates and incubated at 37 °C for 72 h under microaerophilic and anaerobic conditions. Furthermore, its capacity to thrive under various oxygen levels, encompassing microaerophilic and anaerobic conditions, was assessed.
2.2.2 Mupirocin resistance test
Since the isolation procedure involved a medium supplemented with mupirocin, resistance of the isolate to mupirocin was assessed. For comparison, Lactiplacti-bacillus plantarum F75 from the Biology Department culture collection at Universitas Pelita Harapan was used. This strain was originally isolated from the chicken crop. The two bacterial strains were cultured on mMRS agar at 37 °C, under anaerobic and microaerophilic conditions. Following a 72-h incubation time, growth of the strains on agar plates was recorded.
2.2.3 Carbohydrate fermentation test
The isolate ability to ferment various carbohydrates was analyzed using a method described by Dosan et al. [7]. For this test, MRS broth without carbon source was prepared manually by mixing 2.5 g of L-cysteine, 0.05 g of mupirocin, 10 g of peptone, 4 g of yeast extract, 8 g of beef extract, 5 g of NaCl, 2 g of di-ammonium hydrogen citrate, 0.2 g of MgSO4.7H2O, 2 g of FeSO4·7H2O, 0.04 g of MnSO4.7H2O and 1 g of Tween 80 in 1 l of sterile water and mixed. Then, 2% carbohydrate was added to the mixture; this medium was then referred to as MRSP broth. The MRS broth without carbon sources served as the negative control. Carbohydrates included glucose (Merck, USA), fructose (Merck, USA), lactose (Merck, USA), fructo-oligosaccharide (FOS; Xi’an Virgin Biotechnology, China), GOS (Xi’an Virgin Biotechnology, China), inulin (Beneo, Germany) and maltodextrin (Qinhuangdao Lihua Starch, China). To monitor acid production, pH indicator of bromocresol purple (30 mg/L) (Merck, Germany) was added to all media. Isolate was inoculated into MRSP broth and incubated anaerobically at 37 °C for 24 h. The isolate ability to ferment carbohydrates was indicated by a drop in pH, resulting in color change of the medium to yellow. Additionally, the total viable cell count was carried out by serially diluting the liquid culture using 0.1% bacteriological peptone. Then, a 50-μl aliquot of the diluted liquid culture was spread onto MRSP agar plates and incubated at 37 °C for 72 h. Colony forming unit (CFU) was used to assess the bacterial growth in presence of various prebiotics. Results were present as relative growth percentage, with bacterial growth in MRS without carbohydrate serving as the reference point for comparison. Relative growth percentage (%) included ratio of bacterial growth in presence of prebiotics to its growth in absence of prebiotics.
2.3 Assessment of probiotic characteristics
2.3.1 Tolerance to various pH levels and various concentrations of bile salts
Assessment of the isolate tolerance to various pH levels and bile salt concentrations was carried out using a modified method from Khushboo et al. [12]. The isolate was cultured overnight in mTPY broth. Then, a 10% inoculum of the liquid culture was added to mTPY broth adjusted to various pH levels (2, 4, 7 and 9) or various bile salt concentrations (0, 0.5, 1 and 2%). All tubes were incubated anaerobically at 37 °C for 3 h. To assess total viable cell, an aliquot of 50 µl was withdrawn before and after incubation and then spread onto mTPY agar plates. All plates were incubated anaerobically at 37 °C for 72 h.
2.3.2 Antibiotic susceptibility
To assess isolate susceptibility to antibiotics, disk diffusion method was used according to Jorgensen and Turnidge [13]. Isolate was grown anaerobically in mTPY broth at 37 oC overnight. Then, a liquid culture (50 µl) was spread onto mTPY agar plate. A total of 12 commonly used antibiotics from various classes were used in this study (μg per disc): ampicillin, 10; erythromycin, 15; chlorampheni-col, 30; bacitracin, 10; rifampin, 5; vancomycin, 30; neomy-cin, 30; streptomycin, 10; kanamycin, 30; gentamicin, 10; nalidixic acid, 30; and tetracycline, 30 (Liofilchem, Italy). Each disc was transferred onto the surface of the agar plates, which were then incubated anaerobically at 37 oC for 72 h. After incubation, diameter of inhibition zones was recorded. Antibiotic susceptibility of the isolate was categorized based on the zone of inhibition. This categorization was carried out by comparing susceptibility patterns of Lactobacillus spp. and Bifidobacterium spp., as the isolate was initially suspected as Bifidobacterium sp.
2.3.3 Antimicrobial activity
Isolate ability to inhibit growth of pathogenic bacteria was assessed using well diffusion assay [14]. In this study, E. coli and S. aureus from Biotechnology Culture Collection (BTCC) were used. Isolate was cultured anaerobically in TPY broth, while pathogenic bacteria were cultured aerobically in nutrient broth (NB) (Merck, Germany). All tubes were incubated overnight at 37 °C. After incubation, the isolate liquid cultures were centrifuged at 5000× g for 20 min at 4 oC and the resulting cell-free supernatant (CFS) was collected. Liquid cultures of pathogenic bacteria (1%) were mixed with liquefied nutrient agar (NA) (Merck, Germany) in Petri dishes. Upon solidification, wells with a diameter of 6 mm were prepared using sterile pipettes. Then, cell-free supernatant (100 µl) was added into each well and plates were incubated at 37 °C for 24 h. Ampicillin (1 mg/ml) was used as positive control, while TPY broth was used as negative control. After incubation, diameter of inhibition zones was recorded. In well diffusion assay, inhibition zones might be achieved by subtracting diameter of the well from that of the clear area.
2.3.4 Mucosal adhesion ability
To assess mucosal adhesion ability of the isolate, three steps were carried out. First, porcine mucus was purified using modified method from Garcia-Gonzalez et al. [15]. Small intestine of a pig was opened, rinsed with cold PBS and the mucosal layer on the inner lining of the intestine was gently scraped off using spatula. Collected mucus was transferred into 50 ml of cold PBS. The mucosal suspension was centrifuged twice, first at 11,000× g for 10 min and then at 20380× g for 20 min. Supernatant was collected and filtered using cellulose acetate (CA) membrane syringe filters (0.2 μm). Protein concentration in the filtered solution, considered mucin, was estimated using Biuret method. First, bovine serum albumin (BSA) solution was prepared to match the estimated mucin protein concentration. The BSA was filtered using CA membrane syringe filters (0.2 μm). Second, mucin and BSA were immobilized on microplates based on a modified method from Tallon et al. [16]. Mucin and BSA solutions (100 μl per well) were added to various wells of 96-well microplates. Microplates were incubated overnight at 4 °C, then each well was washed twice with 200 μl of PBS. Third, adhesion assay was carried out using the immobilized mucus and BSA on the microplates. The liquid bacterial culture incubated at 37 °C for 16 h was diluted with PBS and three various dilution factors (100 μl each) were selected (N-initial). Diluted liquid cultures were added to each well coated with mucin or BSA and then microplates were incubated at 37 °C for 1, 2 and 3 h. Microplates were washed twice with sterile PBS (200 μl per well) and 200 μl of 0.05% (v/v) Triton X-100 solution were added to the microplates, followed by incubation at 25 °C with agitation at 50 rpm using shaker incubator to allow desorption of adherent bacteria (N-adhere). After 2 h of incubation, N-initial and N-adhere were spread-plated on MRS agar and incubated anaerobically at 37 °C for 48 h. Bacterial colonies on the agar plates were enumerated and the percentage of adhesion cells was calculated using the Eq. 1:
Eq.1
% Adhesion = [log (N-adhere) / log (N-initial)] × 100%
2.3.5 Oxidative stress tolerance
Isolate tolerance to oxidative stress was assessed based on a method from Watanabe et al. [17] with modifications. For this assay, MRS broth without manganese was prepared manually. The composition was as follows: 10 g/l bacteriological peptone, 8 g/l beef extract, 4 g/l yeast extract, 20 g/l D (+) glucose monohydrate, 1 ml of tween 80, 2 g/l diammonium citrate, 5 g of sodium acetate and 0.2 g of magnesium sulfate heptahydrate. MRS broth with manganese was prepared by adding 0.04 g/l manganese (II) sulfate. Isolate was cultured in MRS broth and incubated at 37 °C for 24 h under anaerobic conditions. Then, 1% culture (v/v) was inoculated into two various test tubes, each contained MRS broth with or without manganese. The two test tubes were incubated anaerobically at 37 °C and aliquots of the cultures were sampled upon reaching the exponential and stationary phase. Oxidative stress tolerance was assessed by adding 0.5% (v/v) H2O2 to the cultures. Sampling was carried out within 15–20 min after addition of H2O2. Collected samples were then serially diluted and spread-plated onto MRS agar and then CFUs were counted. The relative abundance (%) [18] included ratio of the cell number after incubation to that before incubation at 100%
- Results and Discussion
3.1 Isolation and identification
A pure isolate from the isolation and purification processes was named SU-KC1a. Morphological observation and biochemical activity tests suggested that SU-KC1a was a rod-shaped, Gram positive, catalase negative, non-acid-fast and non-motile bacterium (Figure 1). All these characteristics were similar to those of Bifidobacterium and Lactiplantibacillus species [19].
Isolation and purification processes were carried out anaerobically to achieve anaerobic lactic acid bacteria such as Bifidobacterium spp. However, molecular identification based on 16S rRNA showed that SU-KC1a was a species of L. plantarum. Since taxonomic nomenclature of L. plantarum species was revised to L. plantarum in April 2020, isolate was named L. plantarum SU-KC1a [20].
Assessing the evolutionary distance of L. plantarum SU-KC1a to other Lactiplantibacillus strains and Bifido-bacterium involved constructing a phylogenetic tree using 16S rRNA sequences of L. plantarum SUKC1a, three L. plantarum isolated from HBM, three L. plantarum isolated from various fermented foods and two Bifidobacterium spp. isolated from HBM as an outgroup (Figure 2). Interestingly, lack of clear separations between L. plantarum isolated from HBM and those from various fermented foods suggested that niche alone might not be an assessing factor in the evolution of Lactiplantibacillus spp. This finding challenged assumption that various niches led to distinct evolutionary paths within the Lactiplantibacillus genus. Variability in microbiota within breast milk, affected by factors such as maternal diet and antibiotic use during pregnancy and lactation, indicates a complex interplay of environmental and genetic factors shaping microbial evolution. This suggests separate evolution of multiple Lactiplantibacillus strains within HBM environments.
As many as 590 various genera in breast milk have been detected through 16S rRNA sequencing, ten most frequently detected genera include Staphylococcus, Streptococcus, Lactobacillus, Pseudomonas, Bifidobacterium, Coryne-bacterium, Enterococcus, Acinetobacter, Rothia and Cuti-bacterium [21,22]. Taghizadeh et al. [23] isolated Lactobacillus spp. from breast milk, including L. gasseri, L. rhamnosus, L. acidophilus, L. plantarum, L. reuteri, L. fermentum, L. animalis, L. brevis, L. helveticus, L. oris, L. casei, L. gastricus, L. vaginalis, L. crispatus, and L. salivarius. To further explain relationships between the niche and evolution of Lactiplantibacillus, additional analyses such as whole-genome comparisons and metabolic profile assessments are recommended. These analyses can provide deeper insights into the genetic and functional differences between Lactiplantibacillus strains originating from various niches, shedding light on their adaptive strategies and evolutionary trajectories.
3.2 Effects of oxygen on cell morphology
Lactobacillus spp. are addressed for their ability to grow in anaerobic and microaerophilic conditions [26]. Based on the findings of this study, L. plantarum SU-KC1a was able to grow without oxygen (anaerobic) and with low oxygen concentrations (microaerophilic). Watanabe et al. [17] reported that L. plantarum could capture oxygen in the presence of manganese metal ions and thus provide aerotolerance characteristics. Moreover, MRS media typically contain manganese metal ions in the form of MnSO4 (0.04 g/l) as a source of nutrition. Figure 3 shows morphology of L. plantarum SU-KC1a cells after anaerobic and microaerophilic incubations. Although cells were rod-shaped, anaerobic incubation resulted in shorter rod shape and smaller cell size. According to Parlindungan et al. [27], stress conditions could cause morphological changes in Lactobacillus spp. as an adaptive response of bacteria to survive. For microaerophilic Lactobacillus spp., anaerobic incubation might cause stress for the cells. To improve survival under stress conditions, rod-shaped LAB formed coccoid-like shapes, which were expected to preserve cell membrane and safeguard internal structures from the external environment, ensuring genetic material intact and preserving low rates of metabolic activities [28]. Morphological changes in L. plantarum SU-KC1a under anaerobic and microaerophilic conditions further emphasized adaptive responses of Lactobacillus spp. to various oxygen environments. However, further studies are needed to assess the effects of oxygen concentration on the growth and stress responses of L. plantarum.
3.3 Mupirocin resistance test
In this study, L. plantarum SU-KC1a could grow on mupirocin selective media (mMRS) anaerobically and microaerophilically. Furthermore, L. plantarum F75 showed no growth on mMRS media (data not shown). This finding suggested that L. plantarum SU-KC1a might possess inherent resistance to mupirocin, indicating potential trait variations within strains of similar species. Pechar et al. [29] reported that mupirocin was often added to the media for isolating bifidobacterial cultures because of the intrinsic resistance characteristics of Bifidobacterium spp. However, it is reported that Lactobacillus spp. are susceptible to mupirocin. Similar to Sunardi et al. [18], L. plantarum SU-KC1a demonstrated high-level resistance to mupirocin that might be acquired by several L. plantarum strains. However, mupA gene, causing high-level mupirocin resistance, was not detected in the genome of L. plantarum SU-KC1a. Therefore, the specific gene responsible for the isolate high-level resistance to mupirocin is still unidentified.
3.4 Carbohydrate fermentation test
- plantarum SU-KC1a demonstrated ability to ferment glucose, fructose, lactose, FOS, GOS, inulin and maltodextrin, as evidenced by the color changes of the media from purplish brown to yellow-brown. Media without carbon sources, serving as a negative control, showed no color changes overall. This alteration in color within the media indicated shifts in pH (Figure 4). Markowiak-Kopec and Slizewska reported that carbohydrate fermentation by Lactobacillus spp. produced organic acids such as lactic acid, acetic acid and short chain fatty acids (SCFA), leading to decreases in pH in the large intestine [30]. Since pH of the media supplemented with various carbohydrates decreased after incubation, these results indicated that L. plantarum SU-KC1a was capable to ferment FOS, GOS, inulin and maltodextrin. Analysis of various responsible enzymes for carbohydrate fermentation has been carried out (data are published soon).
The highest growth of L. plantarum SU-KC1a was observed in media supplemented with GOS, followed by FOS, inulin and maltodextrin. Ability of bacteria to ferment carbon sources can be affected by the degree of polymerization (DP). GOS, as a prebiotic, includes a DP range of 2–7, making it easier to ferment by bacteria. FOS, inulin and maltodextrin include DPs of > 10, 2–60 and 3–11, respectively. Higher DP values might prolong the fermentation duration of FOS, inulin and maltodextrin, compared to GOS fermentation. Bacterial ability to ferment oligosaccharides with a DP > 10 includes half of the speed; at which, oligosaccharides ferment with a DP < 10 [31]. Cao et al. [32] reported that three oligosaccharides (GOS, FOS and MOS/mannooligosaccharides) were able to enhance growth of L. plantarum. In this study, bacterial growth rate was slower when cultured on oligosaccharides rather than glucose. Mandadzhieva et al. [33] reported that most of Lactobacillus strains could ferment mono, di and trisaccharides; however, each strain demonstrated a unique fermentation pattern for various carbohydrate sources. Generally, carbohydrates with lower DP are easier to utilize.
3.5 Tolerance to various pH levels and concentrations of bile salts
To qualify as a probiotic candidate, resistance of cells to various pH levels and bile salt concentrations is critical for their survival in the digestive tract and during the processing of functional foods. Sunardi et al. [18] reported that L. plantarum F75 and L. plantarum SU-KC1a were susceptible to simulated gastric fluid (pH 2) but able to tolerate the fluid after 30 and 60 min of exposure. After 120 min of exposure, L. plantarum SU-KC1a was still detected but at low numbers [18]. Compared to Sunardi et al. [18] who used simulated gastric fluid with pepsin supplementation, this study used MRS broth adjusted to various pH levels. Figure 5A demonstrates that L. plantarum SU-KC1a could preserve its high survival after 3 h of exposure to pH 2. Similar to Huang et al. [34], several L. plantarum strains demonstrated minimal or no decreases in cell viability even after 6 h of exposure to low pH and digestive enzymes (pepsin and trypsin).
A previous study by de Melo Pereira et al. [35] included that bile salt resistance within a concentration range of 0.3–2% was a prerequisite for surviving passage through the human digestive tract. As depicted in Figure 5B, number of L. plantarum SU-KC1a cells decreased with increasing bile salt concentration; however, cells demonstrated robust survival within all concentrations of bile salts. Another study by Sunardi et al. [18] reported that L. plantarum F75 and SU-KC1a were able to survive exposure to 0.3% of bile salts for 3 h. This phenotype was attributed to the capacity of various L. plantarum strains to produce bile acid hydrolases, which catalyzed conversion of conjugated bile salts into free bile salts [36].
3.6 Antibiotic susceptibility
Table 1 shows that L. plantarum SU-KC1a demonstrated resistance to three antibiotics, including vancomycin, streptomycin and nalidixic acid from various classes. These findings were partially similar to those of Sunardi et al. [18] who reported that L. plantarum SU-KC1a was resistant to antibiotics such as vancomycin and nalidixic acid. Antibiotic susceptibility tests in the two studies were carried out using different media: mTPY agar in this study and Mueller-Hinton agar in Sunardi et al. [18]. It is noteworthy that resistance profile of L. plantarum SU-KC1a was not unique, as antibiotic resistances in Lactobacillus spp. to β-lactam, glycopeptides, tetracycline, lincosamides and quinolones (nalidixic acid) have previously been documented. According to Mancino et al. [37], L. plantarum was typically resistant to aminoglycosides (gentamycin, neomycin, streptomycin and kanamycin), bacitracin, quinolone (nalidixic acid) and vancomycin. One of these resistances is intrinsic resistance to Lactobacillus spp., meaning that this resistance type cannot be transferred to other bacteria, suggesting that L. plantarum SU-KC1a may be safe for use as a probiotic. If resistance genes are located on plasmids, there is a risk of horizontal gene transfer to other bacteria [38].
3.7 Antimicrobial activity
Since the beginning of this century, numerous serious bacterial infections have evolved resistance to commonly prescribed antibiotics, creating major healthcare issues worldwide. Hence, there are urgent needs to investigate novel natural sources for antibacterial compounds. Antimicrobial activity of L. plantarum SU-KC1a against E. coli and S. aureus was assessed based on the inhibition zone.Based on the diameter of inhibition zones (Figure 6), L. plantarum SU-KC1a demonstrated greater inhibitions to E. coli (5.0 ± 0.7 mm), compared to S. aureus (2.0 ± 0.7 mm). Malanovic and Lohner [41] reported that antimicrobial activity of Lactiplantibacillus spp. was more visible in Gram-negative bacteria than Gram-positive bacteria. Gram-negative bacteria include outer membranes that are selective for antimicrobials. In this study, concentration of antimicrobial compounds in the cell-free supernatant (CFS) could not be assessed, potentially leading to various results. Further investigations regarding antimicrobial activity can be carried out using broth dilution and time-kill assays [42].
Lactiplantibacillus spp. possess numerous mechanisms for eliminating pathogenic bacteria, including production of inhibitory components such as bacteriocins and lactic acid. Bacteriocins produced by L. plantarum, commonly known as plantaricins, encompass various peptide structures and are generally classified into two main classes. Currently, various types of plantaricin have been identified, including plantaricins A, C, D, E, F, J, K, S, T, Y, 423, 163, 149, 35D, BN, SA6, LC74, KW30, ZJ008, LD1, UG1, NC8, C11 and NA [43]. Plantaricin EF, targets the bacterial membrane, leading to the leakage of intracellular compounds and eventual cell death [44].
They detected that exponential-phase cultures were more susceptible to H2O2, compared to that stationary-phase cultures were. According to Sousa-Lopes et al. [51], cell membrane permeability to H2O2 increased in exponential phase, compared to stationary phase.
- Conclusion
In conclusion, L. plantarum SU-KC1a, originally isolated from HBM in Indonesia, demonstrated significant characteristics as a promising candidate for probiotic uses. This strain demonstrated robust oxidative stress tolerance and antibiotic resistance, particularly to mupirocin, as well as showing ability to thrive with various prebiotic supplementations. Its strong tolerance to acidic conditions and bile salts as well as its significant antimicrobial characteristics and adhesive capabilities to mucin underscore the microbial suitability for diverse environments. These attributes not only highlight its relevance in local contexts, but also suggest its potential as a valuable candidate for probiotics development on a global scale, offering possible benefits for human health and well-being. Further studies are essential to fully realize the microbial therapeutic and commercial potentials.
- Acknowledgements
This study was supported by the Center for Research and Community Development, Universitas Pelita Harapan (grant no. P-078-S/FAST/III/2020 and P-010-S/FaST/V/2021). The authors thank Basic (203) and Advanced (407) Biology Laboratory, Department of Biology, UPH, for providing the necessary facilities.
- Conflict of Interest
The authors report no conflict of interest.
- Authors Contributions
Conceptualization, T.T.J; methodology, M.S. and H.V.; data collection, Y.C., A.F.R, J.O., E.T.P., A.C.J.; writing—original draft preparation, M.S.; writing—review and editing, J.J. and T.T.J. All authors have read and approved final version of the manuscript.
- Bifidobacteria
- Lactobacillus
- Mupirocin
- Prebiotic
- Probiotic
Cómo citar
Citas
Garwolinska D, Namiesnik J, Kot-Wasik A, Hewelt-Belka W. Chemistry of human breast milk: A comprehensive review of the composition and role of milk metabolites in child development. J Agric. Food Chem. 2018; 66(45): 11881-11896. https://doi.org/10.1021/acs.jafc.8b04031
Quin C, Estaki M, Vollman DM, Barnett JA, Gill SK, Gibson DL. Probiotic supplementation and associated infant gut microbiome and health: A cautionary retrospective clinical comparison. Sci Rep. 2018; 8(1): 8283. https://doi.org/10.1038/s41598-018-26423-3
Milani C, Duranti S, Bottacini F, Eoghan C, Turroni F, Mahony J, Belzer C, Palacio SD, Montes SA, Mancabelli L, Lugli GA, Rodriguez JM, BodeL, de Vos W, Gueimonde M, Margolles A, van Sinderen D, Ventura M. The first microbial colonizers of the human gut: Composition, activities and health implications of the infant gut microbiota. Microbiol Mol Biol Rev. 2017; 81(4): e00036-17. https://doi.org/10.1128/MMBR.00036-17
Catenza KF, Donkor KK. Recent approaches for the quantitative analysis of functional oligosaccharides used in the food industry: A review. Food Chem. 2021; 355: 129416. https://doi.org/10.1016/j.foodchem.2021.129416
Shoaib M, Shehzad A, Omar M, Rakha A, Raza H, Sharif HR, Shakeel A, Ansari A, Niazi S. Inulin: Properties, health benefits and food applications. Carbohydr Polym. 2016; 147: 444-454. https://doi.org/10.1016/j.carbpol.2016.04.020
Zhai Z, Yang Y, Wang H, Wang G, Ren F, Li Z, Hao Y. Global transcriptomic analysis of Lactobacillus plantarum CAUH2 in response to hydrogen peroxide stress. Food Microbiol. 2020; 87: 103389. https://doi.org/10.1016/j.fm.2019.103389
Dosan R, Mudana SO, Julyanto CMP, Purnama ET, Sugata M, Jo, Tan TJ. Isolation and identification of Bifidobacterium species from human breast milk and infant feces in Indonesia. Biodiversitas. 2024; 25(2): 337-343. https://doi.org/10.13057/biodiv/d250139
Jia FF, Zhang LJ, Pang XH, Gu XX, Abdelazez A, Liang Y, Sun SR, Meng XC. Complete genome sequence of bacteriocin-producing Lactobacillus plantarum KLDS1.0391, a probiotic strain with gastrointestinal tract resistance and adhesion to the intestinal epithelial cells. Genomics. 2017; 109(5-6): 432–437. https://doi.org/10.1016/j.ygeno.2017.06.008
Nishiyama K, Sugiyama M, Mukai T. Adhesion properties of lactic acid bacteria on intestinal mucin. Microorganisms, 2016; 4(3): 34. https://doi.org/10.3390/microorganisms4030034
Jiang M, Zhang F, Wan C, Xiong Y, Shah NP, Wei H, Tao X. Evaluation of probiotic properties of Lactobacillus plantarum WLPL04 isolated from human breast milk. J Dairy Sci. 2016; 99(3): 1736–1746. https://doi.org/10.3168/jds.2015-10434
Eglash A, Simon L. ABM clinical protocol# 8: Human milk storage information for home use for full-term infants, revised 2017. Breastfeed Med. 2017; 12(7): 390-395.https://doi.org/10.1089/bfm.2017.29047.aje
Khushboo, Karnwal A, Malik T. Characterization and selection of probiotic lactic acid bacteria from different dietary sources for development of functional foods. Front Microbiol. 2023; 14: 1170725.
Jorgensen JH, Turnidge JD. Susceptibility Test Methods: Dilution and Disk Diffusion Methods, In: Jorgensen JH, Carroll KC, Funke G, Pfaller MA, Landry ML, Richter SS, Warnock DW, Richter SS, Patel JB (Eds.), Manual of Clinical Microbiology. 11th Ed. Am Soc Microbiol. 2015: pp. 1253-1273.https://doi.org/10.1128/9781555817381.ch71
City S, Sugata M, Jan TT. Probiotic characterization of Bacillus subtilis SM10.1. J Phys Conf., 2020; 1918: 052025. https://doi.org/10.1088/1742-6596/1918/5/052025
Garcia-Gonzalez N, Prete R, Battista N, Corsetti A. Adhesion properties of food-associated Lactobacillus plantarum strains on human intestinal epithelial cells and modulation of IL-8 release. Front Microbiol. 2018; 9: 2392. https://doi.org/10.3389/fmicb.2018.02392
Tallon R, Arias S, Bressollier P, Urdaci MC. Strain- and matrix-dependent adhesion of Lactobacillus plantarum is mediated by proteinaceous bacterial compounds. J Appl Microbiol. 2007; 102(2): 442-451. https://doi.org/10.1111/j.1365-2672.2006.03086.x
Watanabe M, van der Veen S, Nakajima H, Abee T. Effects of respiration and manganese on oxidative stress resistance of Lactobacillus plantarum WCFS1. Microbiol. 2012; 158: 293-300. https://doi.org/10.1099/mic.0.051250-0
Sunardi J, Purnama ET, Sugata M, Victor H, Tan TJ, Jo J. A comparative assessment of Lactiplantibacillus plantarum isolated from chicken and humans as candidates for probiotics. Biodiversitas. 2023; 24(9): 5198-5206. https://doi.org/10.13057/biodiv/d240964
Mattarelli P, Biavati B, Holzapfel WH, Wood BJB. The Bifidobacteria and Related Organisms: Biology, Taxonomy, Applications. Academic Press, Massachusetts. 2017.
Zheng J, Wittouck S, Salvetti E, Franz CMAP, Harris HMB, Mattarelli P, O’Toole PW, Pot B, Vandamme P, Walter J, Watanabe K, Wuyts S, Felis GE, Ganzle MG, Lebeer S. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901 and union of Lactobacillaceae and Leuconostocaceae. Int J Syst Evol Microbiol. 2020; 70(4): 2782-2858. https://doi.org/10.1099/ijsem.0.004107
Li SW, Watanabe K, Hsu CC, Chao SH, Yang ZH, Lin YJ, Chen CC, Cao YM, Huang HC, Chang CH, Tsai YC. Bacterial composition and diversity in breast milk samples from mothers living in Taiwan and Mainland China. Front Microbiol. 2017; 8: 965. https://doi.org/10.3389/fmicb.2017.00965
Zimmermann P, Curtis N. Breast milk microbiota: A complex microbiome with multiple impacts and conditioning factors. J Infect. 2020; 81(1): 17–47.https://doi.org/10.1016/j.jinf.2020.01.023
Taghizadeh M, Safaei HG, Poursina F. Identification of Lactobacillus plantarum in breast milk. Res Mol Med. 2017; 5(4): 50-60. https://doi.org/10.18502/rmm.v5i4.3065
Tamura K, Nei M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol Biol Evol. 1993; 10: 512-526.
Kumar S. Stecher G, Li M, Knyaz C, Tamura K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol. 2018; 35: 1547-1549. https://doi.org/10.1093/molbev/msy096
Zotta T, Parente E, Ricciardi A. Aerobic metabolism in the genus Lactobacillus: Impact on stress response and potential applications in the food industry. J Appl Microbiol. 2017; 122(4): 857-869.https://doi.org/10.1111/jam.13399
Parlindungan E, Lugli GA, Ventura M, van Sinderen D, Mahony J. Lactic acid bacteria diversity and characterization of probiotic candidates in fermented meats. Foods. 2021; 10: 1019. https://doi.org/10.3390/foods10071519
Parlindungan E, Dekiwadia C, Tran KTM, Jones OAH, May BK. Morphological and ultrastructural changes in Lactobacillus plantarum B21 as an indicator of nutrient stress. LWT. 2018; 92: 556-563. https://doi.org/10.1016/j.lwt.2018.02.072
Pechar R, Rada V, Parafati L, Musilova S, Bunesova V, Vlkova E, Killer J, Mrazek J, Kmet V, Svejstil R. Mupirocin-mucin agar for selective enumeration of Bifidobacterium bifidum. Int J Food Microbiol. 2014; 191: 32-35. https://doi.org/10.1016/j.ijfoodmicro.2014.08.032
Makowiak-Kopec P, Slizewska K. The effect of probiotics on the production of short-chain fatty acids by human intestinal microbiome. Nutr. 2020; 12(4): 1107. https://doi.org/10.3390/nu12041107
Cantu-Jungles TM, Hamaker BR. New view on dietary fiber selection for predictable shifts in gut microbiota. mBio. 2020; 11(1): e02179-19. https://doi.org/10.1128/mBio.02179-19
Cao P, Wu L, Wu Z, Pan D, Zeng X, Guo Y, Lian L. Effects of oligosaccharides on the fermentation properties of Lactobacillus plantarum. J Dairy Sci. 2019; 102: 1-10. https://doi.org/10.3168/jds.2018-15410
Mandadzhieva T, Ignatova-Ivanova T, Kambarev S, Illiev I, Ivanova I. Utilization of different prebiotics by Lactobacillus spp. and Lactococcus spp. Biotechnol Biotechnol Equip. 2014; 25(supl1): 117-120. https://doi.org/10.5504/BBEQ.2011.0132
Huang L, Goda HA, Abdel-Hamis M, Renye Jr JA, Yang P, Huang Z, Zeng Q, Li L. Partial characterization of probiotic lactic acid bacteria isolated from Chinese dairy products. Int J Food Prop. 2021; 24(1): 446–456. https://doi.org/10.1080/10942912.2021.1900233
de Melo Pereira GV, de Oliveira Coelho B, Magalhães Júnior AI, Thomaz-Soccol V, Soccol CR. How to select a probiotic? A review and update of methods and criteria. Biotechnol Adv. 2018; 36(8): 2060-2076. https://doi.org/10.1016/j.biotechadv.2018.09.003
Yang Y, Liu Y, Zhou S, Huang L, Chen Y, Huan H. Bile salt hydrolase can improve Lactobacillus plantarum survival in gastrointestinal tract by enhancing their adhesion ability. FEMS Microbiol Lett. 2019; 366(8): fnz100. https://doi.org/10.1093/femsle/fnz100
Mancino W, Lugil GA, van Sinderen D, Ventura M, Turroni F. Mobilome and resistance reconstruction from genomes belonging to members of the bifidobacterium genus. Microorganisms. 2019; 2019(7): 638. https://doi.org/10.3390/microorganisms7120638
Rozwandowicz M, Brouwer MSM, Fischer J, Wagenaar JA, Gonzalez-Zorn B, Guerra B, Mevius DJ, Hordijk J. Plasmids carrying antimicrobial resistance genes in Enterobacteriaceae. J Antimicrob Chemother. 2018; 73(5): 1121-1137. https://doi.org/10.1093/jac/dkx488
Marbun KT, Sugata M, Purnomo JS, Dikson, Mudana, SO, Tan TJ, Jo, J. Genomic characterization and safety assessment of Bifidobacterium breve BS2- PB3 as functional food. J Microbiol Biotechnol. 2024; 34(4): 871-879. https://doi.org/10.4014/jmb.2311.11031
Campedelli I, Mathur H, Salvetti E, Clarke S, Rea MC, Torriani S, Ross RP, Hill C, O’Toole PW. Genus-wide assessment of antibiotic resistance in Lactobacillus spp. Food Microbiol. 2019; 85(1): 1738-1818. https://doi.org/10.1128/AEM.01738-18
Malanovic N, Lohner K. Gram-positive bacterial cell envelopes: The impact on the activity of antimicrobial peptides. Biochim Biophys Acta. 2016; 1858: 936-946.https://doi.org/10.1016/j.bbamem.2015.11.004
Tanoto J, Sanaky FMN, Sugata M, Tan TJ. Antibacterial activity of Lactobacillus plantarum F75 against Escherichia coli and Staphylococcus aureus. AIP Conf Proc. 2023; 2614: 0020002.https://doi.org/10.1063/5.0127313
Meng F, Zhao H, Zhang C, Lu F, Bie X, Lu Z. Expression of a novel bacteriocin—the plantaricin Pln1 in Escherichia coli and its functional analysis. Protein Expr Purif. 2015; 119: 85-93.https://doi.org/10.1016/j.pep.2015.11.008
Amer SA, Abushady HM, Refay RM, Mailam MA. Enhancement of the antibacterial potential of plantaricin by incorporation into silver nanoparticles. J Genet Eng Biotechnol. 2021; 19(1): 13. https://doi.org/10.1186/s43141-020-00093-z
Kiousi DE, Efstathiou C, Tegopoulos K, Mantzourani I, Alexopoulos A, Plessas S, Kolovos P, Koffa M, Galanis A. Genomic insight into Lacticaseibacillus paracasei SP5, reveals genes and gene clusters of probiotic interest and biotechnological potential. Front Microbiol. 2022; 13: 922689.https://doi.org/10.3389/fmicb.2022.922689
Uraipan S, Brigidi P, Hongpattarakere T. Antagonistic mechanisms of synbiosis between Lactobacillus plantarum CIF17AN2 and green banana starch in the proximal colon model challenged with Salmonella typhimurium. Anaerobe. 2014; 28: 44-53.https://doi.org/10.1016/j.anaerobe.2014.05.002
Prochazkova N, Falony G, Dragsted LO, Licht TR, Raes J, Roager HM. Advancing human gut microbiota research by considering gut transit time. Gut. 2023; 72: 180-191.https://doi.org/10.1136/gutjnl-2022-328166
Mantzourani I, Chondrou P, Bontsidis C, Karolidou K, Terpou A, Alexopoulos A, Bezirtzoglou E, Galanis A, Plessas S. Assessment of the probiotic potential of lactic acid bacteria isolated from kefir grains: Evaluation of adhesion and antiproliferative properties in in vitro experimental systems. Ann Microbiol. 2019; 69: 751–763.https://doi.org/10.1007/s13213-019-01467-6
Mays ZJS, Chappell TC, Nair NU. Quantifying and engineering mucus adhesion of probiotics. ACS Synth Biol. 2020; 9(2): 356-367.https://doi.org/10.1021/acssynbio.9b00356
Dai S, Xie Z, Wang B, Yu N, Zhao J, Zhou Y, Hua Y, Tian B. Dynamic polyphosphate metabolism coordinating with manganese ions defends against oxidative stress in the extreme bacterium Deinococcus radiodurans. Appl Environ Microbiol. 2021; 87(7): 2785-2820.https://doi.org/10.1128/AEM.02785-20
Uhl L, Dukan S. Hydrogen peroxide induced cell death: The major defences relative roles and consequences in E. coli. PLoS ONE. 2016; 11(8): e0159706.https://doi.org/10.1371/journal.pone.0159706
- Resumen ##plugins.themes.ojsPlusA.frontend.article.viewed##: 609 ##plugins.themes.ojsPlusA.frontend.article.times##
- pdf (English) ##plugins.themes.ojsPlusA.frontend.article.downloaded##: 279 ##plugins.themes.ojsPlusA.frontend.article.times##