Synergistic Inhibitory Effects of Nisin and Allicin against Salmonella Typhimurium in Fresh Chicken Meats
食品生物技术的应用,
卷 13 编号 1 (2026),
7 十二月 2025
,
第 1-12 (e10) 页
https://doi.org/10.22037/afb.v13i1.51583
摘要
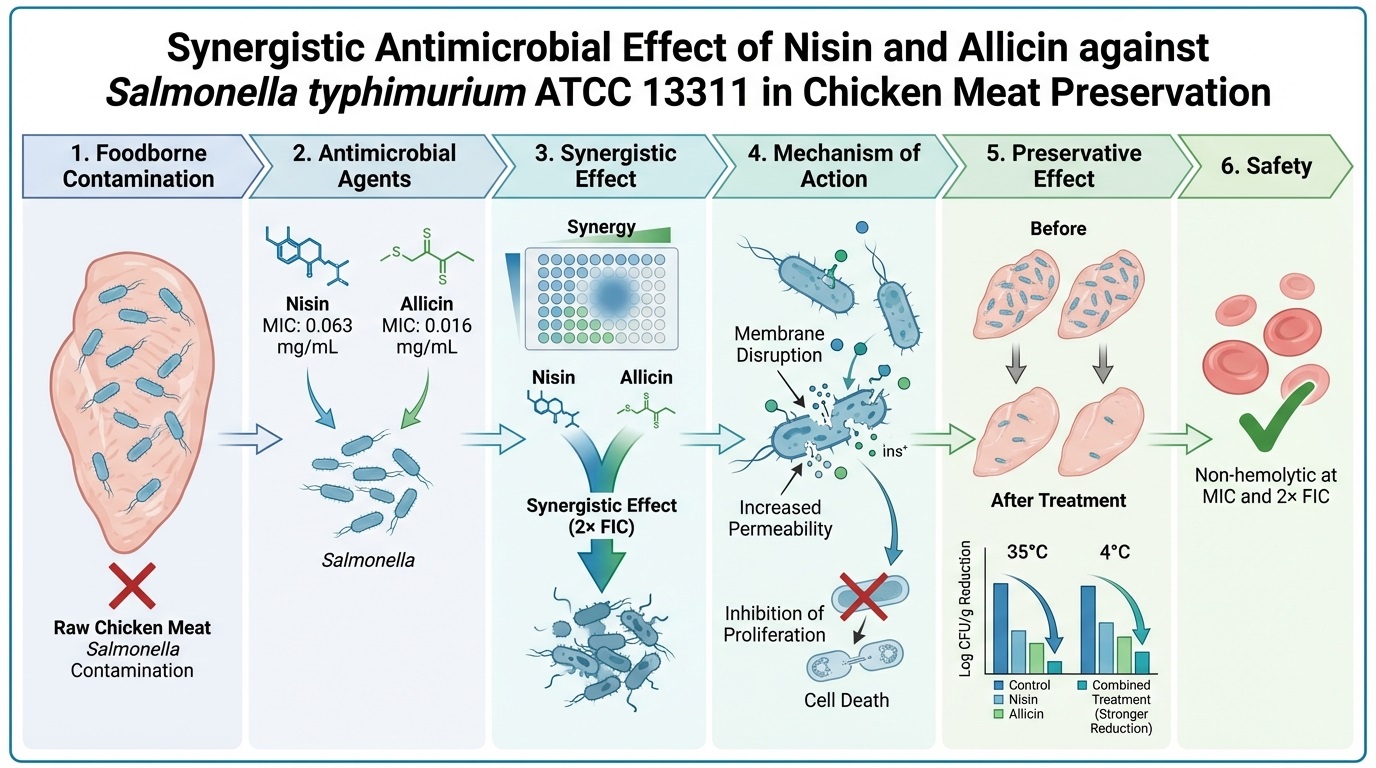
Background and Objective: Foodborne infections represent a significant global health concern, affecting numerous countries. From these, Salmonella Typhimurium ATCC 13311 is a prevalent foodborne pathogen, contaminating poultry products, especially chicken meats. This study investigated the synergistic effect of nisin in combination with allicin against Salmonella Typhimurium ATCC 13311, aiming to enhance the preservation of chicken meats from contamination.
Material and Methods: The inhibitory effect of nisin and allicin against Gram-negative bacterial strains was assessed using microbroth dilution method. Their synergistic effect was investigated through a checkerboard assay. Kinetics of inactivation, membrane permeability using β-galactosidase analysis, hemolytic effect, preservative potentials against Salmonella Typhimurium and calorimetric analysis were assessed.
Results and Conclusion: Nisin and allicin inactivated the proliferation of the test bacteria at minimum inhibitory concentrations of 0.063 and 0.016 mg ml-1, respectively. A combination of the two preservatives was used to treat meat contaminated with Salmonella Typhimurium ATCC 13311. At 35°C, nisin and allicin minimum inhibitory concentrations significantly decreased Salmonella Typhimurium ATCC 13311 colony counts to 2.24 ±0.1 and 2.29 ±0.09 log CFU g-1 after 30 min (p = 0.0193). Moreover, nisin and allicin resulted in 0.08 ±0.02 and 0.09 ±0.02 log CFU g-1, respectively, at 4 °C with no statistical difference compared to the control (p = 0.1064). However, a significant decrease in the population of Salmonella Typhimurium ATCC 13311 at 35 and 4 °C (p = 0.001) was observed when treated with the combination of the two compounds at 2× fractional inhibitory concentration (3.44 ±0.04 and 1.09 ±0.04 log CFU g-1, respectively). Single and combined concentrations were non-hemolytic at minimum inhibitory concentrations and 2× fractional inhibitory concentration. The study suggested the use of nisin and allicin concoctions as a novel approach to control Salmonella Typhimurium ATCC 13311 induced spoilage of chicken meats.
Keywords: Allicin, Chicken meat, Nisin, Synergistic effects, Salmonella Typhimurium
- Introduction
The Food and Agriculture Organization of the United Nations (FAO UN) [1] reported the output of nearly 73 million tons of eggs and poultry meat, exceeding 100 million, 2016. These figures are consistently accelerating due to an increase in the global population, which contributes to rapid economic growth and urbanisation [2]. However, projections for chicken production in 2024 were reported as 146 million tons, an increase of 0.8% over previous years [3]. This increase was due to high consumer demands for chickens, which are affordable despite global inflation. Additionally, developing and developed countries demand poultries and their products [4]. Salmonella foodborne infections account for approximately 85% of the annual global estimates (200 million incidences), with 93% resulting in gastroenteritis and 155,000 fatalities [5]. Moreover, Salmonella infections from chicken and turkey meat consumption cause 23% of the total foodborne infections (1.35 million) in the US, as reported by the Interagency Food Safety Analytics Collaboration [6].
In most food industrial settings, chlorine and organic acids are used in disinfecting materials used in meat processing; however, Salmonella spp. can evade the effect of such chemical interventions, especially when the bacteria produce biofilm [7]. However, biopreservation uses metabolic byproducts and compounds from microorganisms, plants and animals to preserve the sterility of foods while ensuring its safety [8]. Bacteriocins and plant-derived compounds such as nisin and allicin include biopreservative potential against various species of foodborne pathogens. Therefore, they can be used to minimize the challenges faced by food industries when preserving chicken meat against Salmonella spp.
Nisin is one of the purified bacteriocins and accepted as a food additive by the European Union (EU) [9]. A study detected that 0.5% concentration of nisin decreased the number of Listeria monocytogenes to 2.0 log colony-forming units per gram (CFU g-1) in processed turkey ham upon storing at 4 °C for 63 d [10]. Therefore, nisin has been proven as an additive for meat preservation, contributing to the quality and integrity of food in response to consumer demand. Despite the limitation of nisin towards Gram-negative bacteria, its synergy with other natural preservatives can enhance its effectiveness and preservative potential against Gram-negative bacteria, specifically Salmonella Typhimurium, causing chicken meat spoilage [11]. Allicin is a prominent compound of garlic (Allium sativum), with a broad spectrum of activity on various microbial cells using multiple targets, which makes it difficult for bacteria to resist its effect [12]. Allicin can bind to bacterial membrane proteins and phospholipids, enhancing bacterial outer membrane permeability [13]. However, use of allicin alone as a biopreservative may experience unavoidable limitations, especially at high doses, which can cause cytotoxic effects and disrupt the normal gut microbiota [14].
Several studies have reported on the antimicrobial efficacy of nisin on Gram-negative bacteria, including Salmonella Typhimurium [15][16]. Moreover, studies were reported by various scientists on the efficacy of allicin on Gram-negative and Gram-positive bacteria, which were detected effective [17]. However, there is insufficient data on the synergy between the two bioactive agents (nisin and allicin) in inhibition of Salmonella Typhimurium, particularly in fresh chicken meats. These deficiencies created significant gaps for investigating their combined actions in the real food system (chicken meats) and practical context regarding its quality and safety. Although the two compounds have been investigated individually, by verifying their synergistic inhibition against Salmonella Typhimurium under real-life meat storage conditions, the study has provided a better alternative for effective poultry preservation; thereby, decreasing use of synthetic and chemical preservatives in the food industry. Therefore, this study assessed the synergistic antibacterial efficiency of nisin and allicin against Salmonella Typhimurium ATCC 13311 associated with chicken meat contamination under various storage settings.
- Materials and Methods
2.1 Collection and Preparation of the Bacterial Strains
For this research, bacterial strains (Pseudomonas aeruginosa DMST15501, Salmonella Typhimurium ATCC 13311, Escherichia coli ATCC 8239 and Vibrio spp.) were provided from the Microbiology Laboratory, King Mongkut’s University of Technology Thonburi, Thailand. Ten microliters (10 μl) of refrigerated bacterial strains were inoculated onto sterile Luria-Bertani (LB) agar plates and incubated at 37 °C overnight. To achieve a pure culture of the strains, the incubated isolates were re-subcultured and incubated at 37 °C for 24 h.
2.2 Preparation of Nisin and Allicin Stock Solutions
Nisin used in this study was procured from Bangkok Chemical, Thailand, while allicin was purchased from Wang Zhao, China. Nisin and allicin were prepared in mg ml-1. For nisin (2.5% w/w), 10 mg were dissolved in 1 ml of 0.02-N hydrochloric acid (HCl) and set in boiling water (100 °C) for 4 min to enhance its solubility and achieve a uniform solution [15]. However, 2 mg of allicin powder (98% w w-1) were dissolved in 2 ml of 5% dimethyl sulfoxide (DMSO) [18]. The stock solution of nisin and allicin was set at 4 °C before use, with allicin stock wrapped with aluminum foil. The stocks were filtered using 0.22-µm membrane filter (Millipore, USA) to achieve sterility.
2.3 Assessment of Minimum Inhibitory Concentration and Minimum Bactericidal Concentration
To assess the least concentration of the preservatives capable of inhibiting the growth and/or killing the bacterial cells as minimum inhibitory concentration (MIC), a few colonies of the pure isolates were inoculated into tubes containing 1 ml of LB broth and incubated for 2 h. The absorbance of the bacterial suspension was adjusted to 0.5 McFarland level (105 CFU ml-1). After adjustment, 50 μl of the standardized suspension were transferred to the 96 microwells and gently mixed with 50 μl of the prepared concentrations of nisin and allicin, separately. Wells containing LB broth without bacterial suspension were used as negative control, whereas wells containing LB broth and bacterial suspension without preservatives were set as positive control. The analysis was carried out in triplicate, followed by incubation at 37 °C for 18 h. The incubated agar plates were interpreted using microplate reader at 600 nm. The lowest concentration with the same optical density (OD) as that of the positive control was reported as MIC. Ten microliters from the MICs and subsequent concentrations were used to enumerate the MBC by plating onto a labelled sterile nutrient agar and incubating at 37 °C overnight. The concentrations that showed no growth were reported as MBC.
2.4 Inhibitory Synergistic Effects of Nisin and Allicin
The response of the bacterial strains to the combination of nisin and allicin was assessed in the ratio of one-to-one (1:1) as reported by Field et al. [19] with minor modifications. Shortly, fractions of MIC results (1/4) of each preservative were used against the bacterial strains by adopting the checkerboard method. Briefly, 50 μl of the MIC of each preservative were dispensed into a 96-well microplate in vertical and horizontal arrangements. The MICs were further diluted accordingly into fractions as previously stated. The other wells contained the mixture of their corresponding dilutions (25 μl from each preservative). A positive control well was set containing LB and preservatives. The standardized bacterial suspension (105 CFU ml-1) was dispensed into every well, mixed gently and incubated at 37 °C overnight. After incubation, the OD of the combinations was observed using microplate reader, compared to the positive control. The inhibitory synergistic effect of the combination was assessed by calculating their fractional inhibitory concentration index (FICI), as demonstrated by Hossain et al. [20].
FICI = MICN / MICN’ + MICA / MICA’
Where, MICN was the MIC of nisin combined, MICN' was the MIC of nisin alone, MICA was the MIC of allicin combined, and MICA' was the MIC of allicin alone. FICI ≤ 0.5 was synergistic, 0.5-1.0 was additive or partially synergistic, 1.0-4.0 was indifferent and > 4.0 was antagonistic.
2.5 Kinetics of Inactivation
The rate; at which, the cells of Salmonella Typhimurium ATCC 13311 decreased over time upon treatment with nisin and allicin, separately and in combination, was assessed by measuring their OD. A 96-well microplate was used; in which, MICs of nisin, allicin and FICs were dispensed into separate wells in triplicate. A standardized bacterial suspension (105 CFU ml-1) was added to each well. Luria-Bertani broth with bacterial suspension was set as control. The decrease and the increase in their OD were measured using microplate reader at 600 nm every 1 h for 5 h, adopting the procedure of Ajingi et al. [15] with adjustments.
2.6 Beta-galactosidase Analysis as a Model of Membrane Permeability
To assess the major contributing factor toward the synergism of nisin and allicin on E. coli ATCC 8239, its ability to disrupt the bacterial cell membrane (CM) needs assessment. Therefore, this method measured the ability of the β-galactosidase enzyme to leak out of the bacterial cytoplasm into the extracellular space by converting o-nitrophenyl-β-D-galactopyranoside (ONPG) into o-nitrophenol (ONP) [21]. To induce the expression of β-galactosidase, E. coli ATCC 8239 was grown in lactose broth and incubated at 37 °C with shaking (200 rpm) until the OD reached 0.4 at 600 nm. The cells were concentrated by spinning at 5000 rpm for 5 min at 4 °C. The media residue was washed out of the sediment by washing with phosphate-buffered saline (PBS) (pH 7.4). The sediment was diluted in PBS and adjusted to 0.1 OD (107 CFU ml-1) at 600 nm. Fifty microliters (50 µl) of the bacterial suspension were added to 50 µL of MICs of nisin and allicin and their FICs in 96-well plates, followed by 30 µl of ONPG (20 mM). Then, 50 µl of PBS mixed with 50 µl of the bacterial suspension were set as control. The 96-well plate was incubated at 37 °C for 30 min and the OD was read at 405 nm using microplate reader.
2.7 Hemolysis Assay
Red blood cell (RBC) lytic assay was carried out following the procedure explained by Jiddah et al. [21] with minor adjustments. The RBCs were washed in 1× PBS three times and centrifuged at 14,530× g for 10 min. The sediment was dissolved in 1× PBS to achieve a 4% concentration. In general, 500 μl of the RBC dilution were mixed with 500 μl of nisin and allicin, separately and in various combinations (1× FIC and 2× FIC), using tubes. The positive control consisted of a solution with 0.1% Triton X-100, whereas the negative control comprised 1× PBS. The tubes were incubated in a microtube heating block at 37 °C for 1 h and then centrifuged at 14,530× g for 5 min. Then, 100 μl of the supernatant were pipetted from each tube and dispensed into the corresponding wells of the 96-well plate. The release of hemoglobin was assessed by reading the absorbance at 540 nm.
2.8 Inoculation of Chicken Meat Samples
Fresh chicken meat was purchased from the market and washed thoroughly with distilled water (DW) to remove any contaminants. Then, this was cut into pieces (10 g each) using sterilized knife and laminar flow hood. A loopful of fresh overnight culture was transferred to a tube containing 1 ml of LB broth and incubated for 2 h with constant agitation (200 rpm). After incubation, the absorbance of the bacterial suspension was compared with that of the McFarland standard. Nearly 100 μl of the adjusted bacterial suspension (105 CFU ml-1) were distributed on each 10 g of the washed chicken meat samples and allowed bacterial attachment for 30 min before subjecting them to treatment.
2.9 Treatment
The meat samples inoculated with bacterial culture were soaked in 200 ml of the MICs of nisin and allicin and their 2× FIC (0.016 and 0.008 mg ml-1) for 1 h followed by DW as control. Use of 2× FIC was because antimicrobial agents were relatively less active in complex food matrices such as meat due to protein binding, fat content and limited diffusion. This is adopted from the method of Ajingi et al. [15]. After the removal of the meat sample from the solutions, this was drained for 1 h using laminar flow hood. The samples were transferred into sterile polyethylene bags after being treated in two sets. One set was stored at 4 °C using refrigerator and the other set was stored at 35 °C using incubator for 6 d. Samples were analyzed on Days 0, 2, 4 and 6.
2.10 Sampling and Analysis
From the two sets of incubated treated samples (10 g), 90 ml of peptone water were poured into each sample and vigorously smashed for 60 sec to suspend bacteria in the solution using laboratory stomacher. After smashing, the solution was diluted following a tenfold serial dilution in buffered peptone water. After serial dilution, 50 μl of the diluted solution were dispensed into sterile plates, followed by pouring a warm sterilized LB agar solution and incubating at 37 °C for 24 h. After incubation, colonies were counted and recorded in log CFU g-1. In addition to common storage conditions, chicken samples were exposed to 35 °C to mimic severe temperature abuse scenarios that can occur during inappropriate handling, defective equipment and cold chain interruptions. While 35 °C is not appropriate for retail or household storage temperatures, increased temperature was similar to that of recent research, assessing microbial behavior and spoilage under abusive conditions [22].
2.11. Colorimetry Analysis
The sample colors were analyzed using ColorQuest colorimeter (Hunter Associates Laboratory, Inc.,USA). The color values of Hunter (L, a, b), showing lightness, redness and yellowness, were recorded at three sample sites each [15].
2.12 Data Analysis
GraphPad Prism statistical software v.10.4.1 (GraphPad Software, San Diego, CA, USA) was used to analyze data. Statistical differences between the inactivation and membrane permeability were compared using paired t-test. While differences between log decrease and calorimetric parameters were compared using one-way and two-way ANOVA, respectively. The differences were defined as statistically significant (95% confidence interval, p ≤ 0.05).
- Results and Discussion
3.1 Assessment of Minimum Inhibitory Concentration and Minimum Bactericidal Concentration
In this study, a promising alternative for inhibiting the proliferation of Gram-negative bacterial strains, as well as combating the effect of Salmonella Typhimurium ATCC 13311 associated with chicken meat spoilage, was investigated from a globally accepted food preservative and organosulfur compound originating from a natural source. The mean values of the activity of bioactive agents against the Gram-negative bacterial strains are present in Table 1. The results show that nisin and allicin inhibited the proliferation of bacterial strains at various concentrations, with Salmonella Typhimurium ATCC 13311 was the most susceptible, followed by P. aeruginosa DMST15501 and E. coli ATCC 8239, with Vibrio spp. as the least susceptible. The minimum concentrations of nisin and allicin that killed the bacterial strains were higher than their respective MICs. However, Salmonella Typhimurium ATCC 13311 was killed at a lower concentration, compared to the other three bacterial strains. These results suggested that nisin could still inhibit and deactivate Gram-negative bacteria at higher concentrations than Gram-positive bacteria. These findings were supported by studies of Ajingi et al. [15] and Charest et al. [23]. However, the inhibitory and bactericidal effects of allicin in this study was similar to those in another study of Tao et al. [24].
3.2 Inhibitory Synergism of Nisin and Allicin
In this section, FICI was used to assess the synergism of nisin and allicin in all bacterial strains, as shown in Table 2. The FICI value against Salmonella Typhimurium ATCC 13311 was 0.5 when subjected to the combination of 1/4 fraction of nisin and allicin MIC values. In contrast, the concentration used against the other bacterial strains was investigated by how well each preservative acted against that strain. The result revealed that the combination was synergistically effective against all four bacterial strains at various FICs, with FICI less than or equal to 0.5. Moreover, Salmonella Typhimurium was shown as further susceptible to the combination (FIC), including FICI of 0.5.
Nisin, a globally recognized food additive with no allergic reactions or side effects when consumed by humans, includes limitations for activity on Gram-negative bacteria such as Salmonella Typhimurium because of the presence of an outer membrane that delays it from penetrating and attacking CM of the bacteria [25]. Allicin, an active organo-sulfur compound originating from garlic, was reported to improve antimicrobial agents, including peptides and common antibiotics, on Gram-positive and Gram-negative bacteria used for food preservation [26]. Despite the inhibitory potentials of allicin, it can be toxic to eukaryotic cells at a high concentration, which reacts with thiol groups [27]. The synergistic activity of nisin and allicin against Salmonella Typhimurium in this study was similar to that of reports by Ajingi et al. [15] and Duscaron et al. [17]. Additionally, Yang et al. [28] reported synergistic inhibitory effect of nisin and cellulose nanofibrils (CNFs) on various strains of Gram-positive and Gram-negative bacteria with Salmonella Typhimurium.
3.3 Kinetics of Inactivation
The pattern; by which, Salmonella Typhimurium ATCC 13311 inactivated over time when subjected to nisin and allicin at their MICs and FICs was investigated by the fluctuations in their OD at 600 nm for 5 h. The result present in Figure 1 shows that the MICs of nisin and allicin included a moderate inhibitory effect against Salmonella Typhimurium ATCC 13311 through 5 h with a slight increase in their OD, showing incomplete bacterial growth inhibition within the time frame. Moreover, 1× FIC and 2× FIC were more potent, compared to MICs of individual preservatives. Overall, these results suggested that the 2× FIC combination was associated to increased inhibitory activity against Salmonella with a significant difference, compared to other concentrations and the control (p = 0.008), as predicted by synergistic interactions based on the common definition of FICI (≤ 0.5). Patterns of suppression of such a behavior were similar to those of expectations for outcomes that were usually verified through time-kill kinetics, which is still the gold standard to assess antimicrobial interactions [29].
Regarding standardized Clinical Laboratory Standard Institute (CLSI) guidelines for MIC assessment [30], this integration of OD-based evidence established a strong theoretical basis for the efficacy of the nisin and allicin combination (1× and 2× FICs) and highlighted further potential synergistic therapy development. The synergistic inactivation pattern by 1× and 2× FICs in this study was supported by the report of Ajingi et al. [15], when nisin was synergized with organic acid on Gram-positive and Gram-negative bacterial strains.
3.4 β-Galactosidase Analysis
This study investigated the activity of nisin and allicin at their MIC and FIC levels on E. coli ATCC 8239 and linked Gram-negative bacteria such as Salmonella Typhimurium ATCC 13311 by changes in the OD of the treatment after the appearance of yellow coloration, indicating release of the beta-galactosidase enzyme capable of converting ONPG into ONP (Figures 2A and B). The results showed that nisin at its MIC did not cause release of beta-galactosidase, presenting almost a similar OD to that of the control. However, allicin has demonstrated appreciable effects on the bacterial membrane during 60 min after the color change and before the decrease in the OD level. Furthermore, 2× FIC and 1× FIC caused a significant release of beta-galactosidase with a relatively rapid increase in OD level. Additionally, the permeability effect of the FICs was different, with 2× FIC more effective than 1× FIC. Differences in the permeability of FICs (1× and 2× FICs), MICs (nisin and allicin) and control were statistically significant (p = 0.012). This verified synergism between allicin and nisin.
Ideally, nisin inhibits the Gram-positive bacterial cell wall biosynthesis via interruption of trans-glycosylation and mislocalization of lipid II, which is the building block of the bacterial cell wall, leading to the bacterial CM disruption through pore formation [31]. However, nisin alone at lower concentrations faces difficulties to compromise the outer membrane layer of Gram-negative bacteria, as lipopolysaccharide prevents it from contacting with the peptidoglycan layers. Therefore, synergizing nisin with allicin has facilitated its entry into the inner membrane of the bacteria through a gradual process. The combined action of the previously stated mechanisms for nisin and allicin is believed to make their synergy successful against Salmonella Typhimurium ATCC 13311 in this study.
Salmonella Typhimurium is the target pathogen, while E. coli is used as a proxy for β-galactosidase-based permeability assays due to their similarities as Gram-negative Enterobacteriaceae. The two include a double-membrane system with an outer membrane rich in lipopolysaccharides (LPS) [5]. Thus, changes affecting the outer membrane such as increased permeability or antimicrobial disruption similarly affect the two species. Future studies can incorporate Salmonella-specific reporter constructs or stress response markers (e.g., PhoP/PhoQ regulators, oxidative stress indicators and fluorescence-based viability reporters) to provide a pathogen-relevant assay similar to the β-galactosidase assay. These approaches allow mechanistic assessment directly in Salmonella without relying on lacZ-dependent systems.
3.5 Hemolysis Assay
To ascertain the safety of the nisin and allicin at the combined fractional inhibitory concentration used for chicken meat treatment, their hemolytic effect against RBC was studied. Results of the analysis demonstrated that nisin and allicin at their MICs coupled with their double FIC (2× FIC) included no hemolytic activity by showing a small OD at OD540 (similar to the negative control of 1× PBS) with no visible red coloration (Figure 3). However, the positive control (Triton X-100) included a high OD, suggesting that it damaged the RBCs by showing red color after centrifugation. Even though nisin and allicin are bioactive agents originating from natural sources, it was reported that high concentrations and exposure to allicin could begin cytotoxic and erythrocytic oxidative damages to the host cells [31]. Another study reported that allicin might cause dose-dependent hemolysis, supported by phosphatidylserine externalization, calcium influx and oxidative stress [32]. In contrast to allicin, nisin is generally regarded as safe, with limited or no reported cytotoxicity or hemolysis to mammalian cells and RBCs at an antibacterial concentration. Hence, this study has verified the safety of nisin and allicin at the concentration used for the chicken meat treatment; similar to the study of Sarkar and Jayanta [33].
3.6 Inhibitory Effects of Nisin and Allicin on Salmonella Typhimurium ATCC 13311 Contaminating Chicken Meats at 35 and 4 °C Storage Conditions
The synergistic inhibitory effects between the MIC of individual preservatives and their 2× FIC on chicken meats spoilage by Salmonella Typhimurium ATCC 13311 at abusive temperature (35 °C) and refrigerated condition (4 °C) is present in the logarithmic form of their mean values (Log CFU g-1) and standard deviation (Table 3). Colonies of Salmonella Typhimurium ATCC 13311 after 30 min of treatment with nisin, allicin and their FIC combination are present in the table as Day 0. However, the individual preservatives significantly decreased the population of Salmonella Typhimurium ATCC 13311 on Days 2 (0.04 ±0.02), 3 (1.04 ±0.02) and 6 (1.07 ±0.03 and 1.05 ±0.03) with a p-value of 0.0193, compared to the control. Moreover, the 2× FIC caused a more significant difference in log reduction (p = 0.001) of Salmonella Typhimurium ATCC 13311 on the treated meat samples, compared to the control, showing the highest decrease on Day 2 (2.23 ±0.2), 4 (3.21 ±0.1) and 6 (3.15 ±0.03), respectively. Figure 4A illustrates the appearance of the meat sample after treatment with DW and 2× FIC, stored at 35 and 4°C, respectively.
However, allicin showed a significant difference in log decrease of Salmonella Typhimurium ATCC 13311 under refrigerated storage conditions (4 °C) (p = 0.0144). In contrast, no significant difference was seen in the decrease of Salmonella Typhimurium by the MIC of nisin, compared to the control (p = 0.1064) under similar conditions. However, the 2× FIC established a certain preservative effect against Salmonella Typhimurium ATCC 13311 better than that of individual MICs and control, with a significant logarithmic decrease on Days 2 (2.23 ±0.03), 4 (1.24 ±0.04) and 6 (1.18 ±0.04), respectively (p = 0.001).
Despite the ability of Salmonella Typhimurium to adhere to food substances, mostly meats such as chicken and other poultry meats, this study revealed a promising achievement in the decrease of the bacterial growth in fresh chicken meats by the combination of nisin and allicin at their 2× FIC level after 6 d of storage conditions. This effect could be associated with the inhibitory effect of allicin on biofilm formation by Salmonella Typhimurium, which is the major mechanism of the bacteria used to firmly adhere to the meat surface [34]. Mechanisms exerted by allicin on Salmonella Typhimurium are further effective when synergized with other antimicrobials; as reported by Choo et al. [26]. This verified synergism of nisin and allicin against Salmonella Typhimurium ATCC 13311 in the fresh chicken meat samples. The multiplication of Salmonella Typhimurium ATCC 13311 in this study was higher at an abusive temperature (35 °C) than refrigerated conditions (4 °C). The low-temperature effect was possibly further justified because such a low temperature significantly decreased bacterial metabolic processes, which was a factor in minimizing the growth and activity of Salmonella Typhimurium ATCC 13311 during incubation [35]. Additionally, activity of nisin depended on the temperature condition and was reported more active at an increased temperature due to the active state of the target organism, which enhanced the interactions of the nisin and the organism membrane components. However, the log decrease observed after treatment could be associated to allicin augmentation. Although the present study demonstrated antimicrobial effectiveness over the storage, concentrations of residual allicin were not quantified. Further studies incorporating chromatographic techniques such as high-performance liquid chromatography (HPLC) can allow direct assessment of compound stability and degradation kinetics, strengthening long-term efficacy claims.
The synergistic antibacterial effect of nisin and allicin in this study was supported by the findings of Ajingi et al. [15], who reported that nisin in combination with organic acid demonstrated effective preservative potential against Bacillus subtilis in chicken meats and potatoes. Additionally, a study by Singh et al. [36] reported a synergistic effect of nisin and aqueous garlic extract against four strains of L. monocytogenes in a food system stored at 4 °C, similar to the findings of this study. Moreover, the garlic allyl isothiocyanate bioactive compound was reported to enhance the antibacterial activity of nisin against Gram-negative foodborne pathogens, including Salmonella Typhimurium [37]. This verified ability of garlic biocompounds in augmenting other antimicrobials against various species of foodborne pathogens. One limitation of this study included use of 35 °C, as the sample temperature was inconsistent with that commonly used to avoid extreme temperature abuse in chicken meats, which is typically set below 5 °C.
3.7 Calorimetric Analysis
The microbial spoilage of food generally produces undesirable changes in the actual color of the food sample and can be attributed to the secretion of metabolic byproducts, most especially at the stationary phase of bacterial growth within the food sample. In this study, the calorimetric analysis was carried out on the beginning day of the treatment (Day 0) and the final day (Day 6) after storage at 35 and 4 °C, respectively. The results revealed that the chicken meat treated with MICs of nisin and allicin under an abusive temperature demonstrated an increase in the degrees of lightness (L*), redness (a*) and yellowness (b*) over 6 d, with no significant difference with the control treatment (p = 0.2424, 0.2413 and 0.9417, respectively). This suggested higher pigment oxidation and spoilage discoloration at abusive temperatures. The combination of nisin and allicin (2× FIC) helped preserve the natural color of the treated chicken meat samples, leading to a slight increase in lightness (L*) and redness (a*) with significant differences with red color of the control meat sample after 6 d (p = 0.0001). However, the 2× FIC treatment showed no significant difference in the degree of yellowness (b*) with that of the control and the two MIC treatments on the final day of treatment (p = 0.7816).
At the 4 °C refrigeration temperature, the findings for lightness (L*) and redness (a*) were similar to those observed at the higher temperature. Result showed that the lightness of meat did not change significantly between the MIC of nisin and allicin treatments and the control treatments at Day 6 (p = 0.5114). In contrast, the degree of lightness (L*) for their combination (2× FIC) showed a significant difference between the control and MIC treatments (p = 0.001). Similarly, the degree of redness in chicken meat was not significantly affected using either MIC allicin, MIC nisin or the 2× FIC combination at Day 0 (p = 0.168), indicating that these treatments did not interfere with the characteristic red color of the meat during the beginning refrigerated storage (Day 0). However, results demonstrated a significant difference between the combination treatment (2× FIC) and the control at Day 6 (p = 0.0001).
Moreover, MIC treatments showed no significant difference in the yellowness (b*) of the treated meat samples at Days 0 and 6, with a high p-value of 0.114. Meat samples treated with 2× FIC were significantly different from those treated with control (DW) after 6 d of storage (p = 0.0082). This was similar to a study by Charest et al. [23], who stated that nisin contributed to the delay of biochemical processes used by bacterial cells to deteriorate the color of meat samples. Additionally, stability in the color of chicken meat samples stored at 4 °C was linked to the synergism of the combination of nisin and allicin (2× FIC) at chilled conditions; supported by a study of Dacheng Pharma [38]. Results of the calorimetry analysis in this study was correlated to a study by Ajingi et al. (15). Physical appearance of the treated meat samples is provided in Figure 4A. Regarding acceptability and safety of the highlighted synergistic antimicrobials, the current study provided a safer further cost-effective substitute for controlling chicken meats and associated food spoilage caused by Salmonella Typhimurium.
- Conclusion
It was elaborated that nisin included an inhibitory effect against Gram-negative bacteria, when synergized with allicin at lower concentrations. It was also observed that the combination included preservative potentials against Salmonella Typhimurium ATCC 13311, responsible for chicken meat spoilage within 6 d at various temperature conditions. The study indicated that nisin and allicin could synergistically disrupt the CM of Gram-negative bacteria. Furthermore, it was revealed that nisin and allicin included no hemolytic effect on RBCs at their MICs and 2× FIC. The study indicated that the combination of nisin and allicin (2× FIC) used in the treatment of chicken meat samples included no detrimental effect on the color of the treated meats. However, studies are needed to unveil other possible mechanisms investigated by the combination against Salmonella Typhimurium.
- Allicin
- Chicken meat
- Nisin
- Synergistic effects
- Salmonella Typhimurium
##submission.howToCite##
参考
1. Food and Agriculture Organization of the United Nations. The global livestock environmental assessment model. Rome: FAO; 2020. Available from: http://www.fao.org/gleam
2. Mottet A, Tempio G. Global poultry production: current state and future outlook and challenges. World's Poult Sci J. 2017; 73(2): 245–256. https://doi.org/10.1017/S004393391700007
3. Food and Agriculture Organization. Global livestock production and environmental impact: Annual report. Rome: FAO; 2024. Available from: http://avinews.com/en/fao-global-meat-production-to-grow-in-2024-led-by-poultry
4. Mead GC. Processing of poultry. Special Indian Edition/Reprint. Cham: Springer Nature; 2020.
5. Lamichhane BA, Mawad MM, Saleh M. Salmonellosis: An overview of epidemiology, pathogenesis and innovative approaches to mitigate the antimicrobial-resistant infections. Antibiotics. 2024; 13(1): 76. https://doi.org/10.3390/antibiotics13010076
6. Interagency Food Safety Analytics Collaboration. Food illness source attribution estimates for 2019 for Salmonella, Escherichia coli O157, Listeria monocytogenes and Campylobacter using multi-year outbreak surveillance data, United States. U.S. Department of Health and Human Services, CDC, FDA, USDA/FSIS. 2021.
7. Adrianna A, Ewa C, Agata D, Aleksandra B, Wiktoria W, Krzysztof G. Better together-Salmonella biofilm-associated antibiotic resistance. Gut Microbes. 2023; 15:1. https://doi.org/ 10.1080/19490976.2023. 2229937
8. Pisoschi AM, Pop A, Georgescu C, Turcus V, Olah NK, Mathe E. An overview of the natural antimicrobials' role in food. European Journal of Medicinal Chemistry. 2018; 143: 922-935. https://doi.org/10.1016/j.ejmech.2017.11.095
9. Codex Alimentarius. Norma general para los aditivos alimentarios (Codex Stan 192-1995). Codex Alimentarius Commission. 1995.
10. Ruiz A, Williams SK, Djeri N, Hinton JA, & Rodrick GE. Nisin affects the growth of Listeria monocytogenes on ready-to-eat turkey ham stored at four degrees Celsius for sixty-three days. Poultry Science. 2010; 89(2): 353-358. https://doi.org/10.3382/ps.2008-00503
11. Michely BQ, Thaysa FMM, Anielle O, Amarilis SC, Jéssica LM, Odinei HG, Benício AAF, Fernanda VL. Biopreservatives against foodborne bacteria: combined effect of nisin and nanon-capsulated curcumin and co-encapsulation of nisin and curcumin. J Food Sci Technol, 60(2). 2023; 581-589. https://doi.org/10.1007/s13197-022-05641-8
12. Sulin C, Voon KC, Eng HW, Priya M, Sun TT, Phelim V, Chen Y, Pei Pei C. Review on the antimicrobial properties of allicin, both when used alone and in combination with other medications, published in Folia Microbiologica. 2020; 65: 451-465. https://doi.org/10.1007/s12223-020-00786-5
13. Biology Insight Team. Allicin's Antibacterial Mechanisms: Structure and Action. Microbiology. 2025; https://biologyinsights.com/allicin-antibacterial-mechanisms-structure-and-action/
14. Angelos P, Joseph PK, Narelle F, Catherine MB, Geraldine D. Natural anti-microbials for enhanced microbial safety and shelf-life of processed packaged meat. Foods. 2021; 10(7): 1598. https://doi.org/10.3390/foods10071598
15. Ajingi YS, Sudarat R, Jiddah NU, Yuichi K, Nujarin J. Synergistic effect of nisin with acetic and propionic acids inactivates Bacillus subtilis on meat and potato. Biocatal Agric Biotechnol. 2022; 41: 1878-8181. https://doi.org/10.1016/j.bcab.2022.102317
16. Tiejing L, Jin T, Fu H. Study on the Inhibition Effect of Nisin. J Am Sci. 2005; 1(2), 33-37.
17. Duscaron I, Ilic V, Nikolić A, Ćirić M, Soković T, Stanojković T, Kundaković M, Stankovic L, Jubiscaron A, Nikolić L. Cytotoxicity and antimicrobial activity of allicin and its transformation products, Journal of Medicinal Plants Research. 2012; 6(1): 59-65. https://doi.org/10.1100/2012/561823
18. Zainal M, Nurhayati MZ, Indah MA, & Vivi NA. The antimicrobial and antibiofilm properties of allicin against Candida albicans and Staphylococcus aureus - A therapeutic potential for denture stomatitis. Saudi Dental Journal. 2021; 33:105-111. https://DOI.org/10.1016/j.sdentj.2020.01.008 https://doi.org/10.1016/j.sdentj.2020.01.008
19. Field D, Baghou I, Rea M, Gardiner G, Ross R, Hill C. Nisin in combination with cinnamaldehyde and EDTA to control growth of Escherichia coli strains of swine origin. Antibiotics. 2017; 6(4):35. https://doi.org/10.3390/antibiotics6040035
20. Hossain F, Follett P, Vu KD, Harich M, Salmieri S, Lacroix M. Evidence for synergistic activity of plant-derived essential oils against fungal pathogens of food. Food Microbiol. 2016; 53: 24-30. https://doi.org/10.1016/j.phymed.2011.08.060
21. Jiddah NU, Ajingi YS, Rukying N, Rattanarojpong T, Suntornsuk W, Pason P, Jongruja N. Synergistic effects of recombinant AGAAN antimicrobial peptide with organic acid against foodborne pathogens attached to chicken meat. Appl Food Biotechnol. 2024; 11 (1): e21. http://dx.doi.org/10.22037/afb.v11i1.44981
22. Siraj K, Alagapuram K, Gayathri K, Thenmozhi G, Saravanan G. Metagenomics analysis of microbiota in raw chicken stored at different temperatures over storage time in chennai. Indian J Microbiol, 2025; 65(2): 955-960 https://doi.org/10.1007/s12088-024-01299-2
23. Charest AM, Reed E, Bozorgzadeh S, Hernandez L, Getsey NV, Smith L, Galperina A, Beauregard HE, Charest HA, Mitchell M. Nisin Inhibition of gram-negative bacteria. Microorganisms. 2024; 12: 1230. https://doi.org/10.3390/microorganisms12061230
24. Tao Z, Geng D, Tao J, Wang J, Liu S, Wang Q. Synergistic antibacterial effect and mechanism of allicin and an Enterobacter cloacae bacteriophage. Microbiol Spectr. 2023; 11(6): e0315522. https://doi.org/10.1128/spectrum.03155-22
25. Pawar DD, Malik SVS, Bhilegaonkar KN, Barbuddhe SB. Effect of nisin and its combination with sodium chloride on the survival of Listeria monocytogenes added to raw buffalo meat mince. Meat Sci. 2000; 56: 215-219. https://doi.org/10.3390/foods12173273
26. Choo S, Chin VK, Wong EH. Review: Antimicrobial properties of allicin used alone or in combination with other medications. Folia Microbiol (Praha). 2020; 65(3):451-465. https://doi.org/10.1007/s12223-020-00786-5
27. Salehi B, Zucca P, Orhan IE, Azzini E, Adetunji CO, Mohammed SA. Allicin and health: a comprehensive review. Trends Food Sci Technol. 2019; 86:502-516. https://doi.org/10.1016/j.tifs.2019.03.003
28. Yang Y, Liu H, Wu J, Ma J, Lu P. Bio-based antimicrobial packaging from sugarcane bagasse nanocellulose/nisin hybrid films. Int J Biol Macromol. 2020; 161: 627-635. https://doi.org/10.1016/j.ijbiomac.2020.06.081
29. Peleg Micah. Microbial inactivation kinetics models, survival curve shapes and the temporal distributions of the individual Germs' deactivation. Food Eng Rev. 2024; 16: 163-178 https://doi.org/10.1007/s12393-024-09367-5
30. Clinical and Laboratory Standards Institute. Performance standards for antimicrobial susceptibility testing (34th ed., CLSI supplement M100). 2024; Clinical and Laboratory Standards Institute.
31. Chi BK, Huyen NTT, Loi VV, Gruhlke MCH, Schaffer M, Mäder U, Maaß S, Becher D, Bernhardt J, Arbach M, Hamilton CJ, Slusarenko AJ, & Antelmann H. The Disulfide Stress Response and Protein S-thioallylation Caused by Allicin and Diallyl Polysulfanes in Bacillus subtilis as Revealed by Transcriptomics and Proteomics. Antioxidants. 2019; 8(12): 605. https://doi.org/10.3390/antiox8120605
32. Sultan SA, Khawaji MH, Alsughayyir J. Antileukemic activity of sulfoxide nutraceutical allicin against THP-1 cells is associated with premature phosphatidylserine exposure in human erythrocytes. Saudi Journal of Biological Sciences. 2020; 27(12):3376-3384. https://doi.org/10.1016/j.sjbs.2020.09.005
33. Sarkar Payel, Jayanta K, Kundu. Protective role of Allicin on arsenic induced haematological and renal toxicity. Proc Natl Acad Sci, India Sect B Biol Sci. 2023; 95(3): 559-566 https://doi.org/10.1007/s40011-025-01709-5
34. Lihua L, Jianhuit W, Jialini Y, Guanxin L. Effects of allicin on the formation of Pseudomonas aeruginosa biofilm and the production of quorum-sensing controlled virulence factors, Pol J Microbiol. 2013; 62(3): 243-251. https://doi.org/10.33073/pjm-2013-032
35. Jark R, Marches J, Liston. Low-Temperature Growth of Salmonella. J Food Sci. 2006; 33(6): 641-645. https://doi.org/10.1111/j.1365-2621.1968.tb09092.x
36. Singh B, Falahee B, R. Adams Martin. Synergistic inhibition of Listeria monocytogenes by nisin and garlic extract. Food Microbiol. 2001; 18(2): 133-139. https://doi.org/10.1006/fmic.2000.0383
37. Zou Y, Jung LS, Lee SH, Kim S, Cho Y, Ahn J. Enhanced antimicrobial activity of nisin in combination with allyl isothiocyanate against Listeria monocytogenes, Staphylococcus aureus, Salmonella Typhimurium and Shigella boydii. Int J Food Sci Technol. 2013; 48: 324-333. https://doi.org/10.1111/j.1365-2621.2012.03190.x
38. Dacheng Pharma. The antibacterial activity of nisin and its effects. 2025. https://www.dachengpharma.com/news/nisin-and-its-effects/
- 摘要 ##plugins.themes.ojsPlusA.frontend.article.viewed##: 23 ##plugins.themes.ojsPlusA.frontend.article.times##
- pdf (English) ##plugins.themes.ojsPlusA.frontend.article.downloaded##: 16 ##plugins.themes.ojsPlusA.frontend.article.times##