
Functional Characterization of a Novel Heat-stable Recombinant LCI Bacteriocin
食品生物技术的应用,
卷 12 编号 1 (2025),
4 一月 2025
,
第 1-11(e12) 页
https://doi.org/10.22037/afb.v12i1.47824
摘要
Abstract
Background and Objective: Foodborne pathogens represent a substantial threat to living organisms. Therefore, techniques for prolonging food shelf life while ensuring food its quality are imperative practices that must be adopted. Bacteriocins are broadly addressed as preservatives. This study generally characterized LCI peptide as a β-structure antimicrobial peptide and a novel alternative for extending food shelf life.
Material and Methods: The antimicrobial activity of recombinant LCI was assessed against selected Gram-positive and Gram-negative bacterial strains. Temperature, pH and bile salt concentration stability of the antimicrobial peptide were studied. Furthermore, the effect of the peptide on the bacterial membrane was assessed.
Results and Conclusion: The study demonstrated that this novel LCI recombinant bacteriocin included antimicrobial characteristics with wide-spectrum activity against Gram-positive and Gram-negative bacteria. The minimum inhibitory concentrations (MICs) were 50 µg.ml-1 for Micrococcus (M.) luteus ATCC 6633, Staphylococcus (S.) aureus ATCC 6538 and Bacillus (B.) subtilis ATCC 6633 and 100 µg.ml-1 for Gram-negative bacteria when assessed against Escherichia (E.) coli ATCC 8739, Salmonella (S.) typhimurium ATCC 13311 and Vibrio (V.) parahaemolyticus. Time-kill kinetics demonstrated a bactericidal mechanism of action, showing increased antimicrobial efficacy when reported with acetic acid. Membrane permeabilization assessments indicated that LCI created pores in bacterial membranes in a dose-dependent fashion. The peptide stability assessments revealed its heat resistance up to 100 °C for 15 min, while preserving activity in aqueous solutions within pH range of 3–11 and bile salt concentration of 0–2%. These characteristics indicate that LCI may be a viable candidate for antimicrobial uses, especially when used in combination with organic acids.
Keywords: Antimicrobial peptides, Bacteriocins, Broad spectrum activity, LCI, Organic acids
- Introduction
Over 7.8 billion fatalities globally and 56 deaths annually are attributed to foodborne diseases (FBD). These diseases proliferate rapidly due to contaminated foods, with 7.69% of foodborne and foodborne illnesses constituting 7.5% of global losses (56 fatalities) [1]. Pathogens cause foodborne illnesses through toxins and infections at various intervals, generally of bacterial and/or fungal origins. Fruits, vegetables, meats and seafood are often contaminated by various foodborne pathogens. Salmonella Sp., Staphylococcus Sp. Clostridium perfringens, E. coli, Bacillus cereus and, less commonly, Clostridium botulinum (botulism) are the principal foodborne pathogens [2]. Therefore, develop-ment of novel antimicrobial methods in food safety is vital and time sensitive.
Gradisteanu et al., [3], stated that numerous organisms, including insects, plants, reptiles and humans, possessed innate immune systems that use bactericidal peptides as a universal antibacterial strategy. Production of antimicrobial peptides (AMPs) exemplifies this strategy. To maintain self-sustainability and competitive advantages, Gram-positive and Gram-negative bacteria synthesize bactericidal peptides. These peptides are small antimicrobial compounds consisting of 30–60 amino acids (AA). These ribosome-produced peptides show significant variation in size, structure, mechanism of action, spectrum of activity, physicochemical characteristics and target cell receptors. Ghodhbane et al. [4] stated that most bacteria, predo-minantly Gram-positive bacteria and archaea, were believed to synthesize at least one antimicrobial peptide.
A well-known and extensively studied peptide is the bacteriocin, generally produced by the lactic acid bacteria (LAB). Production and release of bacteriocin peptides are included in the most significant probiotic characteristics of the LAB [5]. Bacteriocins protect their producing bacteria by functioning as pore-forming agents, disrupting the cell membranes of target pathogens or inducing other forms of membrane disturbances [6, 7]. According to Niameh et al. [8], biosafety and multiple functionality of bacteriocins, as well as their antimicrobial characteristics, make them appropriate for incorporation into food systems as health promoting components. Furthermore, Verma et al. [9] emphasized the importance of bacteriocins as food preservatives over conventionally chemical methods. Their specificity and biodegradability make them ideal for enhancing shelf life of susceptible food products.
Despite their promising potentials, many AMPs such as bacteriocins are uninvestigated. Another challenge is the high cost of production and decreased efficiency associated with bacteriocin production, which limit their uses [10, 11]. Based on the increasing demands for narrow-spectrum antimicrobial agents against foodborne pathogens, there is a need of continuous identification, functional characteri-zation and use of these AMPs [12, 13]. The LCI peptide is a novel β-structure antimicrobial peptide produced by B. subtilis. Previous studies have reported the high antimic-robial activity of the peptide against pathogens [14, 15]. Furthermore, the peptide is non-toxic and biodegradable with potential uses as natural food preservative as well as feed additive, which can decrease antibiotic use in livestock [16, 17]. However, to enable the practical use of LCI in industries, it is essential to develop production methods that yield sufficient quantities of peptide and provide a clear understanding of its stability. These considerations have prompted interests in investigating this peptide further.
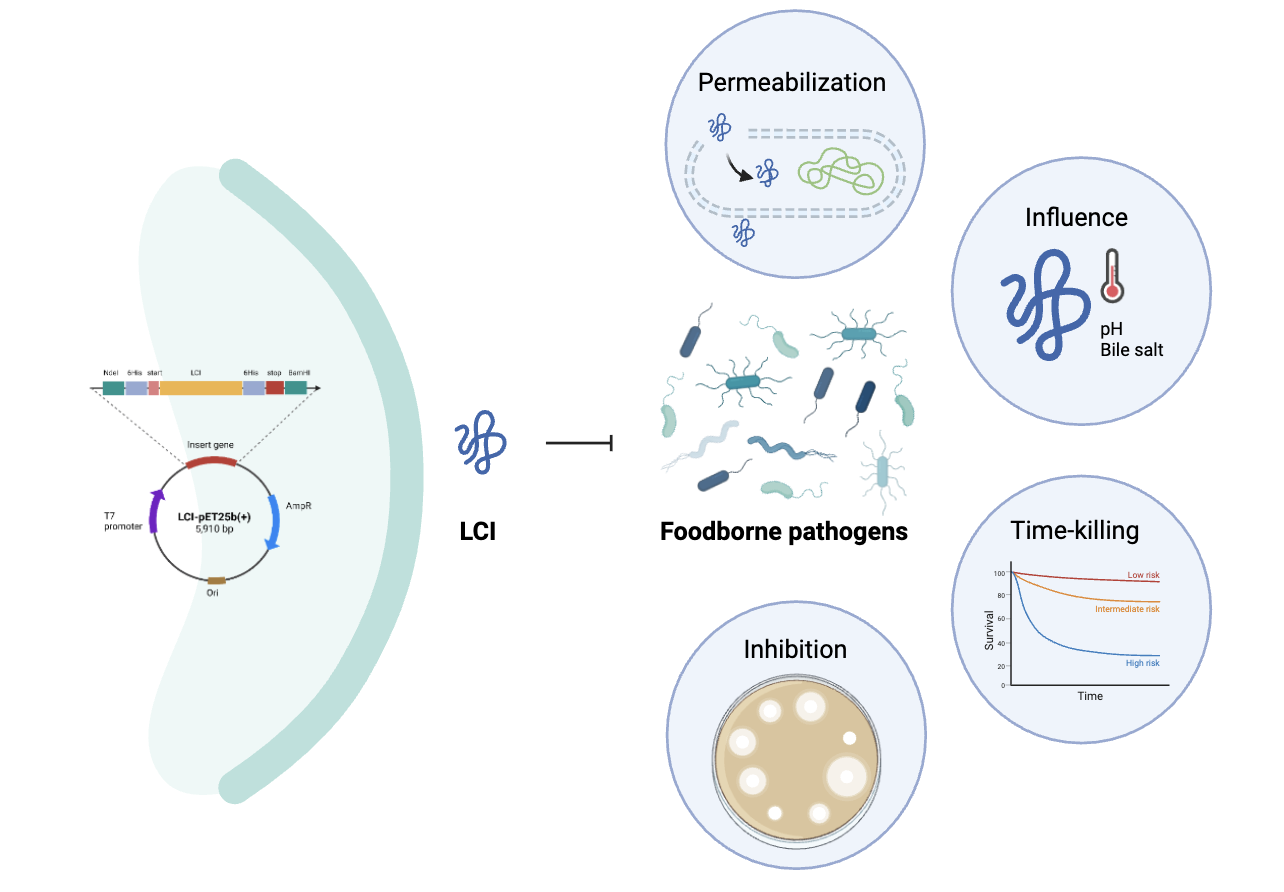
This study investigated the functional characterization of a recombinant LCI peptide derived from the AA sequence of 2B9K.1.A. Antimicrobial characteristics of the LCI peptide were assessed against various Gram-positive and Gram-negative pathogens. Furthermore, the peptide heat resistance, pH tolerance and enzyme sensitivity were assessed. This study advanced antimicrobial peptide research by explaining the structural and functional relationships that caused stability and efficacy of the LCI peptide. Results may direct further studies on peptide-based antimicrobials in microbiology and bioengineering. The peptide shows potentials to enhance public and individual health by improving food safety, decreasing foodborne illnesses and inhibiting antibiotic-resistant microorganisms.
- Materials and Methods
2.1 Bacterial strains, culture conditions, plasmids and oligonucleotides
Expression vectors need multiple components to demonstrate their functions. Humanizing Genomics Macrogen, Bangkok, Thailand, manufactured the codon optimized LCI gene (accession no. APH34379.1) for expression in E. coli. Genomic DNA was amplified using PCR and primers of LCI-F (5'-GGGTTTCATAT-GGCCATTAAACTGGTGCAGTC-3') and LCI-R (5'-GCGGGATCCTCATTAGTGGTGGTGGTG-3'), which targeted the structural gene and incorporate NdeI and BamHI restriction sites, respectively. The PCR products were ligated into NdeI/BamHI-digested pET-25b+ vectors using T4 DNA ligase and then transformed into chemically competent BL21(DE3) E. coli cells at 42 °C for 60 s using heat shock. Colony PCR and sequencing were carried out on transformants to verify integrity of the constructs
2.2 Expression and purification of the recombinant LCI
The pET-25+(LCI) plasmids were transformed into E. coli BL21(DE3) pLysS and cultured in 1 l of 2YT media supplemented with chloramphenicol and ampicillin at 37 °C until optical density (OD) of 0.8 was achieved at 600 nm. Induction of protein expression was carried out by adding 0.1 mM isopropyl-β-D-thiogalactopyranoside (IPTG) to the mixture for 4 h at 25 °C. The bacterial cells were harvested following centrifugation at 5,000× g at for 20 min 4 °C. The cell pellets were collected and stored at -70 °C until use. Cell pellets were resuspended in PBS buffer to 1/5 of the total volume of culture and sonicated for 2 min using microtip connector, pulse duration of 20 s, rest time of 30 s and amplitude of 60%. Then, centrifugation (9,000 rpm at 4 oC for 20 min) was carried out to collect protein in the supernatant. Recombinant LCI was purified using 5 ml HisTrap FF column connected to AKTA prime Fast Protein Liquid Chromatography (FPLC) (General Electric Health-care Systems, USA). The bound recombinant LCI was eluted with 4 ml of elution buffer (10 mM Tris-HCl, 250 mM imidazole and 1 M NaCl; pH 8.0). Then, 16% tricine Sodium Dodecyl Sulfate-Polyacrylamide Gel Electro-phoresis (SDS-PAGE) was carried out to analyze protein fractions and dialysis was carried out at 4 oC overnight using 50 mM Tris-HCl. After protein concentration using centricon tubes (Amicon, Germany), an approximate yield of 0.2 mg of the recombinant LCI was achieved. [18]
Protein concentrations were assessed using Bradford protein assay (Bio-Rad, USA), with bovine serum albumin (BSA) as standard. Purification steps were further assessed on tricine-SDS-PAGE, [19]. Samples diluted in Laemmli buffer [20] were heated at 90°C for 10min before loading onto the gel. Constant voltage of 120V and initial amperage of approximately 50mA were used for 45min. After migration, gels were stained with InstantBlue (Expedeon, UK) for 20min. Liquid chromatography-tandem mass spectrometry (LC-MS/MS) was carried out using Dionex Ultimate 3000 RSLCnano system (Thermo Fisher Scientific, Germany) coupled with an ESI Q-ToF Compact II (Bruker, Germany) to analyze the peptide sequence. Mass spectral data were collected within an m/z range of 200–1,400. Data are present in the supplement.
2.3 Minimum inhibitory concentrations
Minimum inhibitory concentrations (MIC) of the principal bacterial targets, including E. coli ATCC 8739, B. subtilis ATCC 6633, S. aureus ATCC 6538, Salmonella typhimurium ATCC 13311 and V. parahaemolyticus were assessed using broth microdilution assay, following a protocol by Mota-Meira with minor changes. Two-fold serial dilutions of the purified LCI were prepared in 1× PBS and 50 µl of each dilution were dispensed into a 96-well plate. The indicator bacteria cultures grown overnight were diluted to 1 × 10⁸ CFU ml-1 and then 50 µl of them were added to each well containing the purified bacteriocin peptide. Buffer without bacteriocin served as negative control, while those with 100 µg ml-1 bacteriocin served as positive control. The 96-well plate was incubated for 24-h at 37 °C and the OD measured at 600 nm using UT-6550 microplate reader. The MIC was defined as the concentration of bacteriocin that achieved a 50% growth inhibition (MIC₅₀), compared to the positive control. [21, 22] All experiments were carried out in triplicate
2.4 Bacteriocin stability assessment
The peptide antimicrobial activity was assessed using S. aureus ATCC 6538 and E. coli ATCC 8739 as indicator strains. Bacteriocin stability assessments were carried out based on a protocol by Goh and Philip [23]. The LCI peptide samples were incubated at 40, 60 and 80 °C for 40 min and at 100 °C 30 min. The LCI peptide samples were then set to room temperature (RT) prior to assessing their inhibitory effects. The LCI was adjusted to various pH levels using various buffers of 50 mM glycine-NaOH (pH 3–4), 50 mM sodium acetate (pH 4.7), 50 mM tris-HCl (pH 7.9) and 50 mM glycine-HCl (pH 9–11). The inhibitory effect was reassessed after 2 h of incubation at RT. The bacteriocin stability was assessed after exposure to various bile concentrations (0.5, 1 and 2%). Samples were incubated with LCI at 1× the MIC of the reference strains for 2 h at ambient temperature, followed by assessment of inhibitory activity.
2.5 The LCI kinetics of activity
Briefly, S. aureus and E. coli were cultured for 10 h, followed by centrifugation of the media at 2,000 rpm for 5 min to collect the cell pellets. Ice-cold 5 mM sodium phosphate buffer (pH 7.2) was used for the resuspension of each cell pellet. The resuspended cells were mixed at a 1:1 ratio with various treatments of LCI at 1× MIC, 0.03% acetic acid, LCI and 0.03% acetic acid, 1 mg ml-1 nisin and LCI and 1 mg ml-1 nisin and then incubated at 37 °C. A bacterial suspension without LCI served as control. Samples were collected at 20-min intervals to measure the OD600 within a total time of 400 min.
2.6 Assessment of Escherichia coli membrane permeability using beta galactosidase
The E. coli cells were cultivated in lactose broth at 37 °C for 18 h to induce enzyme synthesis. The bacterial cells were harvested by centrifugation to remove the supernatant. The cell pellet was washed three times in 1× PBS buffer and then resuspended and the concentration adjusted to 1 × 10⁶ CFU 100 µl-1 in 1× PBS. Then, 50 µl of the suspension with 50 µl of the purified LCI at concentrations of 2, 1, 0.5 and 0.25× MIC as well as 30 µl of o-nitrophenyl-β-D-galactoside (ONPG) were used as the reaction mixture for membrane permeability assessment. [24] Solutions in 96-well plates were incubated at 37°C for various time intervals. The enzymatic activity was assessed spectrophotometrically at 405 nm.
2.7 Statistic assessment
All experimental results were achieved from triplicate experiments for each sample. The reported data represented the mean of three replicates, using standard deviation (SD) to indicate the level of confidence. Statistical analysis was carried out using GraphPad Prism software (GraphPad, USA). One-way ANOVA was used to assess significant differences between the means (p < 0.05). Error bars represented SDs of the means.
- Results and Discussion
3.1 Synthesis and purification of the recombinant LCI
In preliminary assessments, gene encoding mature LCI was amplified from pCL-LCT and then subcloned into pET-25b+ to create pET-25b+-LCI vector. To enable cleavage of the fusion purification tag (6×His), the expression system was modified. The plasmid construct design used for the LCI expression is shown in Figure 1A. The complete nucleotide sequence of the recombinant plasmid is shown as a chromatogram (Figure 1B). Each peak represented an individual base position in the recombinant plasmid. The sequencing chromatogram showed clear well-resolved peaks, verifying high-quality data with no ambiguous base calls. Comparison of the sequences with the plasmid map of pET-25b (+) containing the bacteriocin gene inserts verified 100% sequence identity of all designed elements. The LCI gene was shown as a single band, with amplicon sizes similar to the theoretical predictions, including approximately 195 bp using LCI primer and 510 bp using T7 promoter primer (Figure 1C).
3.2 Purification of the recombinant LCI and liquid chromatography-tandem mass spectrometry analysis
The low yield of LCI produced by wild B. subtilis A014 was initially a limiting factor that interrupted with the study process. This challenge was overcome by developing a recombinant expression system using host strain of E. coli BL21 (DE3) pLysS to moderate bacteriocin toxicity during background expression. This host could produce within low-temperature expression conditions to better manage protein toxicity and increase peptide concentration [25]. Following IPTG induction, peptides were purified using FPLC. The LCI peptide contained N- and C-terminal His₆-tags. This fusion expression strategy facilitated the purification process. Proteins were eluted using 250-mM imidazole buffer. This approach enabled sufficient peptide production with controlled process and consistent productivity, yielding up to 0.18 mg ml-1. A single elution peak was correlated to antimicrobial activity; as verified in later assessments (Figure 2A). Using this method, 3 mg of the purified LCI were achieved from 15 ml of the soluble fraction. Molecular weight of the purified LCI bacteriocin was assessed as 6.05 kDa using SDS-PAGE. A single protein band was observed after Coomassie brilliant blue staining, verifying the protein purity (Figure 2B).
3.3 Assessment of inhibitory spectra
The purified recombinant LCI specific activity was assessed using serial dilution. The purified LCI bacteriocin was assessed against Gram-positive and Gram-negative bacteria as foodborne pathogens responsible for food poisoning and gastrointestinal diseases in humans. For the five indicator strains, results showed growth inhibition of all bacterial targets as well as significant antimicrobial activities, as indicated by decreases in ODs to less than 50% of the control group (p<0.05), suggesting high susceptibility of the assessed strains. Furthermore, M. luteus ATCC 6633, S. aureus ATCC 6538 and B. subtilis ATCC 6633 showed MIC values of 50 µg ml-1, while Salmonella typhimurium ATCC 13311, E. coli ATCC 8739 and V. parahaemolyticus demonstrated MIC values of 100 µg ml-1 (Table 1). The activity of LCI was similar to that of previously reported pediocin PA-1, which demonstrated efficacy against S. aureus [26]. Moreover, the MIC values indicated that LCI possessed broad-spectrum antibacterial characteristics, advantageous for addressing mixed infections caused by diverse pathogens, similar to bacteriocin Abp118, colicins and sakacin C2 [27–29]. This efficacy is particularly significant as foodborne pathogens pose a persistent threat through microorganisms such as Salmonella typhimurium in the food supply chain, as reported by He et al. [30]
3.4 Kinetics of the recombinant LCI function
Time-kill experiment was carried out to assess the mode of action and rate of inhibitory effects of LCI. The activity of recombinant LCI was assessed by monitoring growth patterns and membrane permeability in S. aureus ATCC 6538 and E. coli ATCC 8739 at various LCI concentrations. The initial mortality rate after incubation indicated that the bacterial growth inhibition depended on the LCI peptide concentration. Results showed that LCI showed a bactericidal mechanism against the target microorganisms, as verified by the sustained decreases in OD within time (Figure 3). After 220 min, the OD at 600 nm decreased significantly from 0.4 to 0.2. This was clearly observed at 100 min, where the Gram-negative bacterial samples treated with acetic acid showed an OD600 of less than 0.5, in contrast to the control and peptide-only groups that respectively showed gradual decreases in ODs after 800 min (Figure 3A) and 150 min for Gram-positive bacteria (Figure 3B), compared to the control group and 0.03% acetic acid group alone. When the peptide is combined with acetic acid, the enhanced antimicrobial efficacy was similar to that previously reported Jiddah et al. [31], who reported enhanced antimicrobial activity of recombinant AGAAN when combined with acetic acid. Similarly, Rothong et al. [32] reported synergistic effects of phage-encoded antimicrobial peptide with organic acid against Acinetobacter baumannii. The increase activity could be resulted due to the disorganization of bacterial membrane, facilitating penetration of the antimicrobial peptide [32].
3.5 Membrane permeabilization assay
This experiment was carried out to assess if LCI exerted its effects on target cells through pore formation. The membrane permeabilization activity demonstrated by β-galactosidase through ONPG assays provided definitive evidence of LCI mechanism of action. The assessment was carried out when the peptide reacted with the ONPG substrate that was leaked out. The OD at 405 nm for untreated bacterial cells was constant (Figure 4). The intensification of yellow color from ONPG and β-galactosidase interaction indicated membrane pore formation. Upon exposure of the reference strain to LCI at concentrations of 0.25, 0.5, 1 and 2× MIC for 600 min, continuous increases in OD measurements were observed, indicating that extracellular o-nitrophenol production significantly increased in E. coli treated with LCI, compared to the control. The experimental findings suggested that this novel peptide analog affected bacterial membrane permeability and permeabilized cytoplasmic membrane of E. coli dose-dependently. Similarly, various bacteriocins such as nisin and plantaricin have similarly demonstrated the ability to disrupt bacterial membranes through pore formation [33–35]. Previous studies [25] reported LCI structural characteristics and suggested a novel mechanism for its action against Gram-negative bacteria. These experimental results verified LCI ability to inhibit Gram-negative bacteria. Additionally, the mode of membrane interaction was similar to 1EWS and MccJ25, which followed a toroidal pore model using positively charged AAs, particularly the AA group at the C-terminal end (Arg46-Lys47). This created short-lived channels in the bacterial membrane through toroidal pore formation, either destroying membrane integrity or penetrating the membrane to affect intracellular targets for antimicrobial effects [36, 37]. The subsequent release of intracellular substances verified the bactericidal characteristics of LCI, facilitating rapid elimination of pathogenic bacteria.
3.6 Effects of temperature, pH and bile salt inhibitors on the bacteriocin
Residual activity was assessed under various temperatures, pH levels and bile salt concentrations to assess LCI stability. The bacteriocin showed stability within a temperature range of 40–100 °C for 15 min. The proportion of surviving bacteria was 31.91% , 47.44% , 46.88% , 65.01% and 74.58% at 4, 40, 60, 80 and 100 °C, respectively (Figure 5A). The findings indicated that LCI included temperature stability, maintaining its complete antibacterial efficacy after 15 min of exposure to increased temperatures. The peptide activity of LCI was assessed under various pH conditions using various buffer systems. Results demonstrated that LCI included various stability and activity depending on the pH of the environment. In 50mM glycine-NaOH buffer, the peptide showed 37.81% and 12.82% activity at pH 3 and pH4, respectively. In 50mM tris-HCl buffer, the activity was 39.34% at pH7 and 42.99% at pH9, indicating relatively higher stability under near neutral conditions to slightly alkaline ones. However, in 50mM glycine-HCl buffer at pH9 and pH11, the peptide activity decreased significantly to 2.48% and 20.93% , respectively. These findings suggested that LCI included greater stability and functionality in tris-HCl buffer at neutral to mildly alkaline pH, while its activity significantly decreased in strongly acidic or highly alkaline glycine-HCl environments (Figure 5B).
Moreover, LCI is characterized by significant stability under various environmental conditions. Its ability to maintain antimicrobial effectiveness at decreased temperatures up to 100 °C within a wide pH range (3–11) makes it an excellent candidate for food processing uses, where such conditions are common. This characteristic was similar to that reported by Johnson et al. [10] and Hols et al., [33], who emphasized that temperature and pH stability were essential for bacteriocins of industrial uses. For comparison, while pH-susceptible agents show optimal activity at neutral pH, bacteriocins such as nisin demonstrate greater efficacy under acidic conditions [34]. To evaluate its potential for use in food and animal feed, the stability of LCI peptide was further tested under gastrointestinal-like conditions. Several antimicrobial peptides lose their effectiveness in bile-rich environments, limiting their uses in gastrointestinal treatments. In the present study, the LCI survivability was assessed in artificial gastric juice at pH 2.5. After 2-h incubation, results indicated high peptide tolerance. The treat with LCI (0% of bile salt) demonstrated the highest antibacterial activity, with a peptide inhibition activity of 69.70±0.006. When exposed to 0.5% bile salt, inhibition efficiency decreased to 49.14±0.008. Similarly, under the condition 1.5% bile salt, inhibition rate was 50.84±0.007. At 2% bile salt concentration, LCI showed moderate activity, with 60.64±0.002 inhibition rate (Figure 5C). These findings indicated that although LCI activity decreased in presence of bile salts, it included significant antibacterial efficacy, particularly at higher bile salt concentrations. In conclusion, LCI demonstrated significant stability at low pH and the peptide preserved its efficacy in presence of bile salts.
- Conclusion
In summary, an effective technique for LCI overproduction in E. coli was established in the present study. A significant quantity of LCI with antibacterial activity was generated through efficient purification. The purified LCI preserved its antibacterial characteristics, demonstrating antimicrobial activity under physiological pH and temperature conditions. The favorable characteristics of LCI suggest its potential as an effective antibacterial therapeutic agent. This study indicates that LCI represents a valuable addition to the growing repertoire of recombinant antibacterial peptides, including potentials to contribute to the development of animal nutrition and serve as a promising alternative to decrease use of antibiotics in the livestock industry.
- Acknowledgements
This study was supported by a research fund from Petchra Pra Jom Klao-PhD scholarship and Faculty of Science Research Fund, King Mongkut’s University of Technology Thonburi, KMUTT.
- Declaration of competing interest
The authors declared no competing interest.
- Authors’ Contributions
- R. Carried out the investigation, writing-original draft, data curation and formal analysis. N. S. Carried out the investigation and data curation. P. D. Carried out the investigation. Y. S. A. Helped in proofreading the manuscript, formal analysis and methodology. N. U. J. Helped in data curation and formal analysis. S. R. Helped in methodology and validation. T. R. Helped in supervision and formal analysis. P. P. Helped in supervision, conceptualization and formal analysis. C. A. Helped in supervision, formal analysis and validation. N. J. Carried out the major supervision, funding acquisition, conceptualization, resources and validation. Findings were discussed by all authors and they contributed to the final manuscript.
- Using Artificial Intelligent Chatbots
No AI chatbot has been used in this study.
- Ethical Consideration
This study did not include any human or animal participants and was carried out in accordance with safety standards.
- Antimicrobial peptides
- Bacteriocins
- Broad spectrum activity
- LCI
- Organic acids
##submission.howToCite##
参考
Lee H, Yoon Y. Etiological agents implicated in foodborne illness worldwide. Food Sci Anim Resour. 2021; 41: 1-7. https://doi.org/10.5851/kosfa.2020.e75
He Y, Wang J, Zhang R, Chen L, Zhang H, Qi X, Chen J. Epidemiology of foodborne diseases caused by Salmonella in Zhejiang Province, China, between 2010 and 2021. Front Public Health. 2023; 11: 1127925. https://doi.org/10.3389/fpubh.2023.1127925
Gradisteanu PG, Popa LI, Marutescu L, Gheorghe I, Popa M, Czobor Bar-bu I, Cristescu R, Chifiriuc MC. Bacteriocins in the era of antibiotic resistance: Rising to the challenge. Pharmaceutics. 2021; 13 (2): 196. https://doi.org/10.3390/pharmaceutics13020196
Ghodhbane H, Elaid S, Sabatier JM, Achour S, Benhmida J, Regaya I. Bacteriocins active against multi-resistant gram-negative bacteria implicated in nosocomial in-fections. Infect Disord Drug Targets. 2015; 15 (1): 2-12. https://doi.org/10.2174/1871526514666140522113337
Allan R, Marcelo BJ, Yvonne LK. Bacterial anti-microbial peptides and nano-sized drug delivery systems: The state of the art toward improved bacteriocins. J Contr Release. 2020; 321: 100-118. https://doi.org/10.1016/j.jconrel.2020.02.001
Etayash H, Azmi S, Dangeti R, Kaur K. Peptide bacteriocins-structure activity relationships. Curr Top Med Chem. 2015; 16: 220-241. https://doi.org/10.2174/1568026615666150812121103
Behrens HM, Six A, Walker D, Kleanthous C. The therapeutic potential of bacteriocins as protein antibiotics. Emerg Top Life Sci. 2017; 1: 65-74. https://doi.org/10.1042/etls20160016
Niamah AK, Al-Sahlany STG, Verma DK, Shukla RM, Patel AR, Tripathy S, Singh S, Baranwal D, Singh AK, Utama GL, Chavez-Gonzalez ML, Alhilfi WAH, Srivastav PP, Aguilar CN. Emerging lactic acid bacteria bacteriocins as anti-cancer and anti-tumor agents for human health. Heliyon. 2024; e37054: 1-15. https://doi.org/10.1016/j.heliyon.2024.e37054
Verma DK, Thakur M, Singh S, Tripathy S, Gupta AK, Baranwal D, Patel AR, Shah N, Utama GL, Niamah AK, Chavez-Gonzalez ML, Gallegos CF, Aguilar CN, Srivastav, PP. Bacteriocins as antimicrobial and preservative agents in food: Biosynthesis, separation and application. Food Biosci. 2022; 46: 1- 23. https://doi.org/10.1016/j.fbio.2022.101594
Johnson EM, Jung DY, Jin DY, Jayabalan DR, Yang DSH, Suh JW. Bacteriocins as food preservatives: Challenges and emerging horizons. Crit Rev Food Sci Nutr. 2018; 58: 2743-2767. https://doi.org/10.1080/10408398.2017.1340870
Abanoz HS, Kunduhoglu B. Antimicrobial activity of a bacteriocin produced by Enterococcus faecalis KT11 against some pathogens and antibiotic-resistant bacteria. Korean J Food Sci Anim Resour. 2018; 38: 1064-1079. https://doi.org/10.5851/kosfa.2018.e40
Sugrue I, Ross RP, Hill C. Bacteriocin diversity, function, discovery and application as antimicrobials. Nat Rev Microbiol. 2024; 22: 556-571. https://doi.org/10.1038/s41579-024-01045-x
Xiaoli C, Hong L, Shuangping L, Jian M. Impact of bacteriocins on multidrug‐resistant bacteria and their application in aquaculture disease prevention and control. Rev Aquac. 2024; 16: 1286-1307. https://doi.org/10.1111/raq.12897
Chen S, Zhao Z, Qian L, Meng Q, Peng L. A novel class I bacteriocin LCI shows strong activity against Gram-positive bacteria. Biochem Biophys Res Commun. 2020; 523(2): 405-411. https://doi.org/10.1016/j.bbrc.2019.12.081
Zheng W, Zhang Z, Li D, Tang J, Xu H. Stability and activity of bacteriocins in various environmental conditions. Life. 2023; 13(7): 1521. https://doi.org/10.3390/life13071521
García-Bayona L, Comstock LE. Bacteriocins in the era of antimicrobial resistance. Front Cell Infect Microbiol. 2022; 12: 876147. https://doi.org/10.3389/fcimb.2022.876147
Yang SC, Lin CH, Sung CT, Fang JY. Antimicrobial peptides: Functional activity and potential applications in food preservation. Front. Microbiol. 2019; 10 (57): 1-17. https://doi.org/10.3389/fmicb.2019.00057
Madi-Moussa D, Coucheney F, Drider D. Expression of five class II bacteriocins with activity against Escherichia coli in Lacticaseibacillus paracasei CNCM I-5369, and in a heterologous host. Biotechnol Rep. 2021; 30: 1- 8. https://doi.org/10.1016/j.btre.2021.e00632
Schägger H, Von Jagow G. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal Biochem. 1987; 166: 368-379. https://doi.org/10.1016/0003-2697(87)90587-2
Laemmli HK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970; 227: 680-685. https://doi.org/10.1038/227680a0
Zhang Y, Yang Y, Shi L, Cao Z, Yang J, Wang J. Purification, characterization, and mode of action of a novel bacteriocin BM173 from Lactobacillus crustorum MN047 and its effect on biofilm formation of Escherichia coli and Staphylococcus aureus. J Dairy Sci. 2020; 103(12): 10665-10676. https://doi.org/10.3168/jds.2020-18506
Al-fekaiki DF, Niamah AK, Al-Sahlany STG. Extraction and identification of essential oil from Cinnamomum zeylanicum barks and study the antibacterial activity. J Microbiol Biotechnol Food Sci. 2017; 7(3): 312- 316. http://dx.doi.org/10.15414/jmbfs.2017/18.7.7.312-316
Goh HF, Philip K. Purification and characterization of bacteriocin produced by Weissella confusa A3 of dairy origin. PLoS ONE. 2015; 10(11): e0140434. https://doi.org/10.1371/journal.pone.0140434
Ajingi YS, Rukying N, Jiddah NU, Koga Y, Jongruja N. Cloning, recombinant expression, purification, and functional characterization of AGAAN antibacterial peptide. 3 Biotech. 2023; 13(3): 88. https://doi.org/10.1007/s13205-023-03512-3
Gong W, Wang J, Chen Z, Xia B, Lu G. Solution structure of LCI, a novel antimicrobial peptide from Bacillus subtilis. Biochemistry. 2011; 50 (18): 3621-3627. https://doi.org/10.1021/bi200123w
Song DF, Li X, Zhang YH, Zhu MY, Gu Q. Mutational analysis of positively charged residues in the N-Terminal region of the class IIa bacteriocin pediocin PA-1. Lett Appl Microbiol. 2014; 376: 356-361. https://doi.org/10.1007/s12274-021-4015-6
Hahn-Löbmann S, Stephan A, Schulz S, Schneider T, Shaverskyi A, Tusé D, Giritch A, Gleba Y. Colicins and Salmocins-new classes of plant-made non-antibiotic food antibacterials. Front Plant Sci. 2019; 10: 437. https://doi.org/10.3389/fpls.2019.00437
Corr SC, Li Y, Riedel CU, O’Toole PW, Hill C, Gahan CG. Bacteriocin production as a mechanism for the anti-infective activity of Lactobacillus salivarius UCC118. Proc Natl Acad Sci. 2007; 104: 7617-7621. https://doi.org/10.1073/pnas.0700440104
Gao Y, Li D, Sheng Y, Liu X. Mode of action of Sakacin C2 against Escherichia coli. Food Control. 2011; 22: 657-661. https://doi.org/10.1016/j.foodcont.2010.07.010
Yue HE, Jikai W, Ronghua Z, Lili C, Hexiang Z, Xiaojung Q, Jiang C. Epidemiology of foodborne diseases caused by Salmonella in Zhejiang province, China, between 2010 and 2021. Front public health. 2023; 11: 1127925. https://doi.org/10.3389/fpubh.2023.1127925
Jiddah NU, Ajingi YS, Rukying N, Rattanarojpong T, Suntornsuk W, Pason P, Jongruja N. Synergistic effects of recombinant AGAAN antimicrobial peptide with organic acid against foodborne pathogens attached to chicken meat. Appl Food Biotechnol. 2024; 11 (1): e21. http://dx.doi.org/10.22037/afb.v11i1.44981
Rothong P, Leungtongkam U, Khongfak S, Homkaew C, Samathi S, Tandhavanant S, Ngoenkam J, Vitta A, Thanwisai A, Sitthisak S. Antimicrobial activity and synergistic effect of phage-encoded antimicrobial peptides with colistin and outer membrane permeabilizing agents against Acinetobacter baumannii. Peer J. 2024; 12: 1-19. https://doi.org/10.7717/peerj.18722
Hols P, Ledesma-García L, Gabant P, Mignolet J. Mobilization of microbiota commensals and their bacteriocins for therapeutics. Trends Microbiol. 2019; 27: 690-702. https://doi.org/10.1016/j.tim.2019.03.007
Prudêncio CV, dos Santos MT, Vanetti MCD. Strategies for the use of bacteriocins in gram-negative bacteria: Relevance in food microbiology. J Food Sci Technol. 2015; 52: 5408-5417. https://doi.org/10.1007/s13197-014-1666-2
Gradisteanu PG, Popa LI, Marutescu L, Gheorghe I, Popa M, Czobor BI, Cristescu R, Chifiriuc MC. Bacteriocins in the era of antibiotic resistance: Rising to the challenge. Pharmaceutics. 2021; 13: 196. https://doi.org/10.3390/pharmaceutics13020196
Vincent PA, Morero RD. The structure and biological aspects of peptide antibiotic microcin J25. Curr Med Chem. 2009; 16: (5), 538-549. https://doi.org/10.2174/092986709787458461
Dobson A, Cotter PD, Ross RP, Hill C. Bacteriocin production: A probiotic trait? Appl Environ Microbiol. 2012; 78: 1-6. https://doi.org/10.1128/aem.05576-11
- 摘要 ##plugins.themes.ojsPlusA.frontend.article.viewed##: 485 ##plugins.themes.ojsPlusA.frontend.article.times##
- pdf (English) ##plugins.themes.ojsPlusA.frontend.article.downloaded##: 169 ##plugins.themes.ojsPlusA.frontend.article.times##