
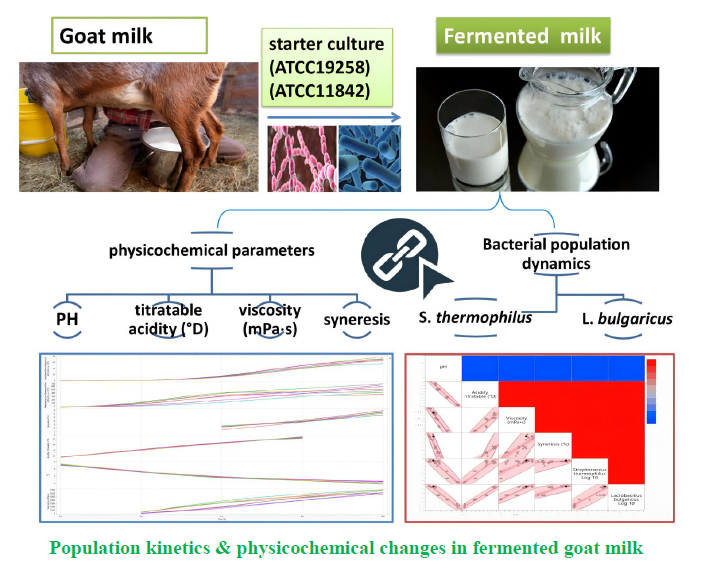
Bacterial Population Kinetics and Physicochemical Profiles in Fermented Goat Milks: Roles of Streptococcus thermophiles ATCC19258 and Lactobacillus bulgaricus ATCC11842
食品生物技术的应用,
卷 12 编号 1 (2025),
4 一月 2025
,
第 1-12 (e7) 页
https://doi.org/10.22037/afb.v12i1.47532
摘要
Abstract
Background and Objective: The fermentation of Algerian goat milk, a process for the production of valuable dairy products, relies on the synergistic activity of Streptococcus (S.) thermophilus and Lactobacillus (L.) bulgaricus. However, a significant knowledge gap is seen regarding the precise dynamics of these starter cultures within the unique matrix of Algerian goat milks. Specifically, the intricate relationships between their growth patterns and the resulting physicochemical changes, which regulate the distinct biochemical characteristics of fermented products, are poorly understood. So, this study addressed this problem by studying specific contributions of S. thermophilus and L. bulgaricus to goat milk fermentation.
Material and Methods: Goat milk was fermented by starter cultures of S. thermophilus and L. bulgaricus (8 h). Bacterial growth and physicochemical parameters, including pH, titratable acidity, viscosity and syneresis, were assessed. Mixed-effects models were used for statistical analysis to assess the relationship between physicochemical changes and bacterial growth.
Results and Conclusion: The results showed a strong relationship between L. bulgaricus and the control of acidification, viscosity and syneresis (r = 0.979 for titratable acidity, p < 0.0001). S. thermophilus contributed significantly, particularly to the increases in viscosity (r = 0.773, p < 0.01). The two species significantly decreased the pH, with L. bulgaricus having twice the acidifying effects. By the end of the fermentation process, pH reached 4.12 ±0.20, titratable acidity increased to 84.75 ±2.19 °D and viscosity increased to 6425.00 mPa.s ±638.64. The final bacterial counts of S. thermophilus and L. bulgaricus were 519.00 ±115.29×10⁷ and 65.54±6.89×10⁷ CFU.ml-1, respect-ively. In addition to providing a robust statistical framework for process control and quality assurance in fermented milk manufacture, this study highlighted the critical role of L. bulgaricus in regulating structural and sensory qualities of fermented goat milks. Results can be used to optimize fermentation processes for goat milk by strategically manipulating the ratio of L. bulgaricus to S. thermophilus. The strong correlation between L. bulgaricus and acidification, viscosity and syneresis (r = 0.979 for titratable acidity, p<0.0001) provides a clear target for controlling key product attributes.
Conflict of interest: The authors declare no conflict of interest.
- Introduction
In Algeria, where goat farming is an essential part of the rural economy particularly in the dry and semi-arid regions with an estimated 4.2 million goats, goat milk processing into fermented products offers significant advantages. This process meets the growing consumer demand for quality-processed foods while optimizing the use of a readily available but currently underused resource. Because of their synergistic effects on the physicochemical and sensory characteristics of fermented dairy products, Streptococcus (S.) thermophilus and Lactobacillus (L.) bulgaricus are freq-uently used as starter cultures in cow milk fermentation. In this study, these strains were used in the fermentation of goat milk, which is novel for facilitating investment in the industrialization of this milk and guaranteeing its organo-leptic and nutritional quality. This focus is critical, especial-ly considering that factors such as milking frequency have been shown to significantly affect the nutritional and microbiological quality of cow milk in Algeria [1]. However, the precise dynamics of these cultures in goat milk, particularly the distinct biochemical characteristics of Algerian goat milk, are not fully understood. These bio-chemical features can directly affect fermentation kinetics and finished product characteristics, such as a higher concentration of short and medium-chain fatty acids (SCFA and MCFA, respectively). In addition, studies on conven-tional dairy production systems have demonstrated the importance of endogenous strains in regulating the unique qualities of local products [2]. These microorganisms contribute to gel formation, improved viscosity, acidific-ation and modification of sensory qualities such as flavor and texture [3].
The use of probiotics to improve metabolic health is a relatively novel indication for probiotic therapy. The poten-tial for probiotics to modulate inflammatory status is particularly interesting, as demonstrated in cell cultures [4]. These two strains show a valuable synergistic relationship, stimulating the other strain growth through the exchange of metabolites in a process known as protocooperation [5]. Specifically, L. bulgaricus expresses extracellular protease to use milk proteins, providing an abundant nitrogen source for itself and S. thermophilus is described to supply L. bulgaricus with certain acids (e.g. formic and folic acids) and carbon dioxide [6]. Additionally, S. thermophilus synth-esizes several amino acids and expresses a cell envelope proteins [5]. The synergistic effect between L. bulgar-icus and S. thermophilus accelerates milk fermentation and enhance microbial growth. Therefore, the primary objective was to investigate the milk fermentation process with particular emphasis on the interactions between S. thermo-philus and L. bulgaricus. A key question is if each bacterium significantly contributes to physicochemical changes in milk, specifically acidification, viscosity and syneresis, critical factors in product innovation. While bacteria such as Streptococcus, Lactobacillus and Bifidobacterium Sp. have extensively been studied in various milk types, research on their interactions in goat milk is limited despite the unique characteristics of goat milks [7].
Fermented dairy products, particularly those derived from goat milks, are significant components of traditional and modern diets, offering valuable nutritional and probiotic benefits. The fermentation process, driven by bacterial cultures such as S. thermophilus and L. bulgaricus, involves a complex interaction of biological and physicochemical factors. Understanding the dynamics of these bacterial populations and their interactions with their surrounding environment is critical for ensuring consistent product quality and safety. Existing research has identified key physicochemical parameters affecting bacterial growth during fermentation, including temperature, pH and substrate availability. However, a significant gap is seen in quantifying these interactions. While qualitative observations are abundant, lack of robust predictive models that can accurately describe the relationships between bacterial growth kinetics and these parameters, particularly within the unique matrix of Algerian goat milks, is addressed. This limitation delays the precise control and optimization of industrial fermentation processes. Furthermore, the specific population kinetics of S. thermophilus and L. bulgaricus in Algerian goat milks, regarding its unique composition affected by local breeds and environmental factors, requires in-depth investigation. The current knowledge may not fully capture dynamics of these bacteria in this specific context. The lack of understanding can lead to variability in product qualities, inconsistent fermentation outcomes and safety concerns.
This study addressed gaps in understanding of lactic acid fermentation in Algerian goat milks. It was beyond general explanations by providing a statistically rigorous analysis that quantified specific roles of S. thermophilus and L. bulgaricus. Unlike previous studies that often treated starter cultures as single entities, the present study highlighted the individual contributions of each bacterium to acidification, viscosity and syneresis. Mixed-effects models were used to establish strong correlations between bacterial growth and physicochemical variations. This study on Algerian goat milk challenged the conventional emphasis on cow milk in fermentation research. This alternative substrate offers potential advantages for consumers pursuing less allergenic dairies. The experimental approach reveals the development of distinct microbial strains and flavors associated with L. bulgaricus. Advanced statistical modelling combined with detailed bioanalysis and industrial uses represents a significant advancement in the current knowledge. Practical recommendations emerging from the results can optimize local transformation processes and promote sustainable goat farming, effectively bridging the gap between fundamental research and real-world effects.
- Materials and Methods
The raw milk used in the experiment was purchased from local goats in a dairy farm in Tissemsilt, Algeria. This was stored at 4 °C before fermentation. The L. bulgaricus ATCC 11842 and S. thermophilus ATCC 19258 were purchased from a specialized food biotechnology supplier (Fly Chemicals). To avoid contamination, these cultures were used to inoculate the milk at a concentration of 10⁷ CFU.ml-1 under aseptic conditions in the laboratory. Raw milk was inoculated with L. bulgaricus and S. thermophilus at a concentration of 10⁷ CFU.ml-1, respectively, and then incubated at 42 °C for 8 h. Samples were collected every 2 h to assess pH, acidity, viscosity and syneresis.
- Physicochemical analyses
The pH was assessed regularly using calibrated pH meter (Hanna HI-2211, Romania) based on AFNOR standards. The acidity was assessed using titration with 0.1 N NaOH and ISO 6091:2010 method [8]. Acidity was expressed in Dornic degrees (°D), where 1 °D included 0.1 g of lactic acid in 100 ml of milk. The viscosity was assessed using viscometer (Fungilab, Alpha series, Spain). Syneresis was assessed using centrifuge (Sigma D-37520, 3-18KS, Germany). The syneresis rate was the percentage of whey separation with the total volume of the product and centrifuged at 1,125 and 3,125 g.
- Microbiological Analyses
The microbiological analyses aimed to quantify the number of lactic acid bacteria (LAB) and pathogens. For the bacterial enumeration, samples were inoculated onto MRS (Man, Rogosa, Sharpe) agar and incubated at 37 °C for 48 h. Results were expressed as colony-forming units (CFU.ml-1).
- Identification of Pathogenic Bacteria
Potential pathogens, including Escherichia coli and Listeria monocytogenes, were detected using ISO [9] method for E. coli and ISO 11290-1, 2017[10] method for L monocytogenes.
- Data Analysis
Data analysis was carried out using JMP Pro 17 software. Bacterial concentrations were log₁₀-transformed to normal-ize the distributions and stabilize the variance. The initial concentrations of S. thermophilus and L. bulgaricus ranged 0.56–682 × 10⁷ and 0.19–594 × 10⁷ CFU.ml-1, respectively. Linear mixed-effects models were used to analyze the dependent variables (pH, titratable acidity, viscosity and syneresis), considering fixed effects of the transformed concentrations and the random effects of time measure-ments. Although indications suggested nonlinear relation-ships, a linear model was chosen to avoid overfitting the data. The model validity was verified using several diagnostic procedures such as tests for the normality of residuals and assessment of homoscedasticity. The AIC and BIC criteria were used to assess model fit and R² statistics were used to assess the model explanatory power. These analyses verified the robustness of the model in assessing effects of lactic acid fermentation on the physicochemical characteristics of raw milks.
- Results and Discussion
During the early fermentation phase (2h), pH decreased to 5.61 ±0.05, whereas the titratable acidity increased significantly to 40.25 °D ±1.58. The first measurable viscosity readings were recorded as 171.63 mPa⋅s ±49.32 (120–270 mPa⋅s). Bacterial populations showed early growth, with S. thermophilus increasing to 6.41 ±8.33 × 10⁷ CFU.ml-1 and L. bulgaricus reaching 1.73 ±0.05 × 10⁷ CFU.ml-1 (Table 1). By mid-fermentation (4 h), significant changes were observed in all parameters (Table 1). The pH decreased to 4.77 ±0.10, accompanied by increased titratable acidity (66.75 °D ±1.75). Initial syneresis was observed (4.78% ±0.71) and the viscosity increased substantially to 2562.50 mPa⋅s ±1263.71. The S. thermo-philus showed exponential growth, reaching 386.50 ±166.36 × 10⁷ CFU.ml-1, whereas L. bulgaricus increased to 5.72 ±0.13 × 10⁷ CFU.ml-1. Within 6 h (Table 1), fermentation progressed with the pH decreasing to 4.50 ±0.02 and the titratable acidity increasing to 84.75 D°±2.19. Syneresis increased to 7.50% ±0.51 and the viscosity reached 4876.25 mPa⋅s ±708.88. The S. thermo-philus showed a slight decrease to 361.38 ±85.41 × 10⁷ CFU.ml-1, whereas L. bulgaricus showed continues growth, reaching 51.58 ±6.04 × 10⁷ CFU.ml-1. At the end of fermen-tation (8 h), the samples reached their lowest pH (4.12±0.20, ranging 3.83–4.32) with maximum syneresis (12.75% ±0.88) and viscosity (6425.00 mPa⋅s ±638.64). The final bacterial counts showed that S. thermophilus and L. bulgaricus increased to 519.00 ±115.29 × 10⁷ and 65.54 ±6.89 × 10⁷ CFU.ml-1, respectively (Table 1).
3.1. Bacterial Population Dynamics
Critical shifts and variations were observed in the population patterns of L. bulgaricus and S. thermophilus over the fermentation time. Similar to the findings of Moghadam et al. S. thermophiles and L. bulgaricus experienced significant growth during fermentation [11]. This phase of growth is critical for acidification. Data indicated that the L. bulgaricus population reached 1.73 ±0.05 × 10⁷ CFU.ml-1, while S. thermophilus increased to 6.41 ±8.33 × 10⁷ over the first 2 h [12]. This advanced phase, in which S. thermophiles increased in quantity, boosted deacidification of the liquid media. The study highlighted the need of S. thermophilus to help acidification process while allowing the remaining environment appropriate for L. bulgaricus to thrive. The growth patterns of the two species differed at 4-h point. During this time, the population of S. thermophilus increased significantly, reaching 386.50 ±166.36 × 10⁷ CFU.ml-1.
In contrast, the population of L. bulgaricus increased, showing a modest increase to 5.72 ±0.13 × 10⁷ CFU.ml-1. Moreover, S. thermophilus showed a stable population at and after the 6-h point, while the L. bulgaricus count increased, achieving a maximum 65.54 ±6.89 × 10⁷ CFU.ml-1 at 8-h time point. These findings were similar to those by Meng et al., who detected synergistic interspecific cooperation between the two bacterial species through omics approaches[13]. This verified that S. thermophiles initiated the fermentation process, whereas L. bulgaricus continued to thrive during the later stages of fermentation. Previously, Ahsan et al. (2022) have shown a broader range of food matrices; to which, these bacteria could be adapted. They reported that S. thermophilus and L. bulgaricus could be used in the fermentation of soy milks [14]. As stated, this study illustrated the changes in population sizes at various stages of fermentation and those at the final stage. The authors previously verified the established claims that the two strains included a synergistic relationship; in which, S. thermophilus was the first to initiate acid production for improving conditions of L. bulgaricus growth. Supporting evidence has further been clarified at the molecular level.
Correlation analysis revealed intricate relationships between bacterial growth and the physicochemical parameters during fermentation (Fig. 1). The pH demon-strated significant negative correlations with all assessed variables (p < 0.01), showing particularly strong negative relationships with titratable acidity (r = -0.982) and L. bulgaricus growth (r = -0.958). This underscored the fundamental role of pH in modulating the fermentation environment. The L. bulgaricus showed significantly stronger correlations with physicochemical parameters than those S. thermophiles did, suggesting its predominant effects on the product characteristics. Specifically, L. bulgaricus showed strong positive correlations with titratable acidity (r = 0.979) and viscosity (r = 0.929) while maintaining a strong positive relationship with S. thermophiles growth (r = 0.898). In contrast, S. thermo-philus showed a moderately strong correlation with viscosity (r = 0.773) and a significantly weaker correlation with syneresis (r = 0.405), indicating its secondary role in texture development.
The textural characteristics demonstrated distinct correlation patterns, with viscosity showing stronger associations with L. bulgaricus (r = 0.929) than S. thermophilus (r = 0.773). Syneresis demonstrated a similar issue, correlating stronger with L. bulgaricus (r = 0.798) than S. thermophilus (r = 0.405). These relationships suggested that L. bulgaricus played a further important role in texture development, possibly through enhanced exopolysaccharide production and/or proteolytic activity. The strong correlation between bacterial growth and physicochemical parameters was further validated by non-parametric analyses (Kendall tau-B and Spearman rho), verifying the robustness of these relationships regardless of the statistical approach used. These findings correlated with those by Nadirova and Sinyavskiy (2023), who reported that L. bulgaricus could grow despite decreased pH levels, owing to its intrinsic adaptability to acidic conditions[15].
Data collected at 6 and 8-h intervals further illustrated the growth patterns of these bacteria. At 6-h point, population of S. thermophiles slightly stabilized, measuring 361.38 ±85.41×10⁷ CFU.ml-1. In contrast, L. bulgaricus demonstrated a further vigorous growth rate, reaching a population of 51.58 ±6.04 × 10⁷ CFU.ml-1. At 8-h point, S. thermophiles population showed slight stability, with an increased count of 519.00 ±115.29 × 10⁷ CFU.ml-1, while L. bulgaricus demonstrated strong growth, achieving a population of 65.54 ±6.89 × 10⁷ CFU.ml-1. The present results have been verified in other products such as yoghurt, where S. thermophilus and L. delbrueckii subsp. bulgaricus multiplied significantly during fermentation [16].
These findings addressed those of Sieuwerts et al. (2010), whose transcriptome analysis revealed that mixed-culture growth involved upregulation of biosynthesis pathways for nucleotides and amino acids that were vital for the growth of the two bacteria [17]. Furthermore, Nadirova and Sinyavskiy (2023) highlighted the complementary roles of these two bacterial species in the fermentation process. The S. thermophilus was essential in initiating acidification, whereas L. bulgaricus proliferated in acidic environments created by the other strain. This complex interaction not only enhanced fermentation efficiency but also contributed to development of the desired flavor and texture profiles in fermented dairy products. In contrast to the findings of Picon et al. (2016), the present findings demonstrated that acidification occurred at a slower further gradual rate [18]. This was primarily attributed to the diverse range of naturally occurring LAB strains that affected lactic acid production efficiency. Conversely, optimized interactions between S. thermophilus and L. bulgaricus strains led to further rapid acidification, increased lactic acid production and consistent acidification profile.
3.2. PH
The linear mixed-effects model analysis of bacterial fermentation dynamics revealed a significant effect of bacterial growth on pH regulation during fermentation (Fig. 2). This model, showing excellent fit characteristics (AICc = -6.299, BIC = -0.686), verified a baseline pH of 6.087 ±0.058 (SE, p < 0.0001) in goat milks. A significant rapid acidification of milk with pH decreasing from 6.62 ±0.08 to 4.12 ±0.20 over 8 h. This pH decrease, critical for flavor development and microbial stability, was primarily driven by the action of the two bacterial species with L. bulgaricus showing a stronger effect (-0.589 ±0.083 pH units per log₁₀ CFU.ml-1, p< 0.0001), compared to S. thermophiles (-0.304 ±0.060 pH units per log₁₀ CFU.ml-1, p<0.0001). This approximately two-fold difference in acidification capacity was statistically supported by highly significant F-ratios for S. thermophiles (F₁,₃₇ = 25.66, p<0.0001) and L. bulgaricus (F₁,₃₇ = 49.74, p < 0.0001), demonstrating their differential contributions to fermentation. The model random effects structure indicated negligible temporal variances, sugges-ting consistent acidification patterns over time. The residual variance was small (0.043 ±0.010), supporting the model precision and reliability. A significant negative correlation (r = -0.898) between the effects of S. thermophiles and L. bulgaricus underscored the complex interactive dynamics between these species rather than a simple additive effect on pH decrease. These findings were similar to those of previous studies, which identified L. bulgaricus as a primary driver of acidification in milk fermentation systems [7,19,20]. Similarly, another study showed that S. thermo-philus promoted rapid bacterial growth and enhances fermentation efficiency, creating a favourable environment for L. bulgaricus. This synergistic relationship led to improved metabolite profiles, with S. thermophilus facilit-ating the production of flavor compounds during milk fermentation at optimal temperatures [19]. A recent study using omics analyses has revealed the molecular mechanisms driving bacterial synergy, driving decreases in pH [12]. This intricate relationship between bacterial inter-actions and pH dynamics was further supported by Wu et al., whose study linked bacterial population ratios and fermentation times to the sensory profile, including acidity [19]. This revealed the broader implications of these pH changes, as they resulted from bacterial metabolic activity and affected the final quality and sensory characteristics of the product. Thus, bacterial populations strongly affected pH dynamics, particularly L. bulgaricus, which showed a greater acidifying capacity, verifying its key roles in the acidification process.
3.3. Titratable Acidity
A linear mixed-effects model analyzed the relationship between bacterial populations and titratable acidity (Fig. 3). The model demonstrated satisfactory fit indices (-2 residual log likelihood = 159.71, AICc = 172.84, BIC = 177.22). Fixed-effects analysis demonstrated highly significant positive relationships between the titratable acidity and logarithmic populations of the bacterial species. The L. bulgaricus showed a stronger effect (β =21.44 ±1.47 D.log₁₀ CFU-1, t (29) = 14.55, p<0.0001), compared to S. thermophiles (β = 7.43 ±0.97 °D.log₁₀ CFU-1, t (29) = 7.67, p<0.0001). This approximately three-fold difference in effect size indicated that L. bulgaricus was the primary driver of acid production in the fermentation system. The model intercept of 30.20 ±0.92 °D (t (29) = 32.85, p<0.0001) represented the baseline acidity when controlling for bacterial populations.
The significance of these relationships was further supported by fixed-effects tests, which showed strong evidence of the effect ofS. thermophilus[F(1,29)=58.83, p<0.0001] and L. bulgaricus [F (1, 29) = 211.58, p<0.0001]. The substantially larger F-statistic for L. bulgaricus verified its dominant role in acid production, with direct implications for starter culture formulations in fermented dairy products. The random effects structure analysis revealed that the time component was confounded with residual variance, resulting in a residual variance estimate of 10.60 ±2.78 °D (95% CI: 6.72–19.16 °D). This confounding factor suggested that bacterial population dynamics, rather than time-dependent factors, were the primary determinants of acid development in this system, which is an important consideration for process control in industrial settings.
Model diagnostics supported the validity of these statistical assumptions. The actual by predicted plots demonstrated a strong linear relationship between the observed and predicted values across the full range of measurements (10–90 °D). Residual analysis revealed a generally symmetric distribution near zero (-6 to +6 °D), with the residual quantile plot indicating approximate normality. The model strong predictive capability suggested that it could be a reliable tool for controlling acidification processes in fermented dairy production.
While goat milk (19.05 °D) and cow milk (17 °D) showed distinct initial acidity levels, the titratable acidity significantly varied during the fermentation process [20]. A significant increase was observed, rising from 15.88 °D ±0.64 to 84.75 °D ±2.19 within 8 h. This increase strongly correlated with bacterial growth, particularly that of L. bulgaricus, whose metabolic activity significantly contributed to milk acidification through lactic acid production. These verified the findings that the proliferation of L. bulgaricus directly affects titratable acidity and that lactic acid production is essential for controlling final characteristics of the fermented product[21,22]. Andrew et al. emphasized that titratable acidity is a relevant indicator of the progression of fermentation process in L. bulgaricus based products. The results of Abbasalizadeh et al. indicated that the maximum lactic acid production in the Media12 media reached 35.01 g.l-1) [23]. This result verified the current results as the titratable acidity reached 84.75 °D ±2.19 after 6 h of fermentation. These results demonstrated the importance of acidification in ferment-ation processes.
Further supporting the present results, Sonnier et al. showed the synergistic actions of S. thermophilus and L. bulgaricus that promoted rapid acidification of the media [24]. More specifically, Qiu et al. provided mechanistic insights into the current findings by revealing the metabolic pathways and metabolites involved in acid production, particularly the role of lactic acid production by L. bulgaricus. According to Wu et al., the link between bacterial ratios and acidity strengthened the connection to product sensory attributes and verified that specific bacterial ratios affected acidity and sensory profile. The present study highlighted the strong acidifying capacity of L. bulgaricus, which was twice that of S. thermophilus. Although the study on L. plantarum SU-KC1a did not directly assess acid production in fermentation, it demonstrated robust tolerance to pH variations [21]. This suggested that L. plantarum SU-KC1a might contribute to acidification during fermentation, although not as strongly as L. bulgaricus. These findings underscored the role of L. bulgaricus as the primary contributor to acidity and the importance of understanding dynamics of bacterial populations for process control, while reinforcing these conclusions by contextualizing them within the current understanding of these mechanisms and their effects on organoleptic characteristics of the final product.
3.4. Viscosity
Analysis of the fixed effects revealed a significant difference in the effect of the two bacterial species on viscosity development (Fig.4), with L. bulgaricus demonstrating dominant effects and coefficient of 3106.28 ±397.51 mPa⋅s per log unit increase in cell density (p < 0.0001), approximately 8.84 times greater than the effect observed for S. thermophilus (351.37 ±281.73 mPa⋅s per log unit, p = 0.2223). This difference demonstrated that L. bulgaricus largely explained viscosity of the fermented media. The random effects structure revealed negligible temporal variance [t (h) = 0], indicating that viscosity changes were not significantly affected by the duration of the experiment. The residual variance was significant (881,027.22), indicating that variations in viscosity were predominantly explained by the bacterial concentrations rather than temporal patterns. Model diagnostics supported these findings; the actual-by-predicted plot demonstrated a generally linear relationship with increased variability at higher viscosity levels (0–7000 mPa⋅s). The model strong statistical characteristics (F-ratio for L. bulgaricus= 61.06, p<0.0001) provided robust evidence for the differential effects of these bacterial species on viscosity development. However, several considerations guaranteed further attentions. First, the negative intercept (-731.81 mPa⋅s) represented a theoretical value outside biologically relevant conditions and should be interpreted cautiously. Further-more, although the model demonstrated heteroscedasticity (unequal variance of the residuals) at higher viscosity values, this did not invalidate the primary findings regarding the relative effect of each species. Of the changes in viscosity, the most significant was observed after 8 h when the viscosity reached 6425.00 mPa.s ±638.64, which was more than two times higher than the initial value. These results were similar to those of Qiu et al., who identified L. bulgaricus with its primary activity of exopolysaccharides (EPS) production as the major reason for the improvement in viscosity. In other words, data were similar to those that addressed L. bulgaricus as an essential component in boosting the product viscosity, predominantly through the production of EPS. A highly positive association (r = 0.929) was detected between the abundance of L. bulgaricus and the magnitude of viscosity, which provided further evidence for this microbial leading role in improving the rheological characteristics of the final product. The importance of this finding is that viscosity is a critical characteristic that dictated the degree of product acceptance; thus, texture and mouthfeel were directly affected by viscosity. The present results were similar to those of Nadirova and Sinyavskiy, who underlined that the increase in viscosity was necessary not only for improving the texture and stability of fermented products during storage but also for providing favorable conditions for the growth of relevant bacteria. This is a further step and a part of bacteriophage resistance of these isolates. Moreover, a project by Afzal et al. on the specific structure of the EPS produced by L. bulgaricus and how researchers verified that the produced EPS could make a difference in the viscosity levels of the final product of fermented milk were significantly verified [25]. Through the interactions of L. bulgaricus EPS with indigenous strains or natural ingredients, viscosity and texture could be optimized; thereby, extending the shelf life of quality products.
syneresis in fermented dairy products (Fig. 5). The L. bulgaricus demonstrated a strong positive association with syneresis (β = 5.26 ±0.92, p < 0.0001), indicating that higher concentrations of this strain significantly increased water expulsion from the gel matrix. While S. thermophilus showed a positive development (β = 3.67 ±2.49, p = 0.1561), its larger standard error (SE) and non-significant p-value suggested significant variability in its effects on syneresis. The intercept of the model (β = -8.68 ±6.23, p = 0.1786) was not significantly different from zero, suggesting minimal baseline syneresis in the absence of bacterial activity. Model diagnostics supported the validity of the present findings, with residual analyses showing appropriate distribution patterns and no substantial violations of the model assumptions. The significantly positive coefficient for L. bulgaricus was strong across multiple diagnostic assessments, reinforcing its critical role in controlling syneresis. However, significant residual variances and wide confidence intervals for some parameters suggested that additional factors such as protein concentration, pH dynamics and temperature fluctuations might contribute to syneresis variation in ways that were not captured by this model.
The fermentation phase was highlighted by increases in syneresis of 4.78% ±0.71 and a significant separation of lactoserum from the gels with syneresis increasing to as high as 12.75% ±0.88. The correlation coefficient between L. bulgaricus and syneresis (r = 0.798) was confusing with that presented by Nadirova, who argued that L. bulgaricus might improve gel syneresis and increase gel strength. This is an important question. Although (EPS) are known to originate from L. bulgaricus and alter gel structures favorably. Two studies showed that specific EPS structures antagonized syneresis, highlighting that L. bulgaricus produced EPS that improved water retention [26]. One interpretation of the present results is that L. bulgaricus under specific fermentation conditions changes the balance of syneresis positively. Certain conditions of the present experiment and a certain strain of L. bulgaricus used in this experiment may need further investigations. However, it is possible that high concentrations of L. bulgaricus and its byproducts facili-tated whey removal and gel retention was compromised. Qiu et al. reported that while the EPS synthesize by L. bulgaricus helped lessen syneresis, the effect was incomplete [12].
The large residual variances and broad ranges of confidence intervals in the present model reveal that various other components such as protein concentrations, pH shifts and temperature changes, which were not seen in the present model, might include effects on syneresis. Overall, the study provided evidence of a further complex relationship between L. bulgaricus, EPS formation and syneresis and results indicated needs of further integrated understanding to enhance the fermentation process. Additional studies are needed to investigate which specific strains lead to decreased syneresis under what conditions, modelling and Optimization.
During fermentation, the population dynamics of S. thermophilus and L. bulgaricus showed complex significant fluctuations (Table 2). At early stages, the two bacterial species showed exponential growth, aligning with the findings of[15], which highlighted the significant growth of S. thermophilus and L. bulgaricus during fermentation. This rapid growth is critical for acidification. The present data indicated that within the first 2 h of fermentation, the population of S. thermophilus increased to 6.41 ±8.33 × 10⁷ CFU.ml-1, while L. bulgaricus reached 1.73 ±0.05 × 10⁷ CFU.ml-1. This initial phase, highlighted by the swift growth of S. thermophilus, was vital for the starting of the acidification of liquid media, similar to other findings by Qiu et al. Their study emphasized the significant role of S. thermophilus in initiating acidification; thereby, creating a further favorable environment for the growth of L. bulgaricus. After 4 h, growth rates of the two species were shifted. At this point, population of S. thermophilus significantly increased to 386.50 ±166.36 × 10⁷ CFU.ml-1, while L. bulgaricus showed a further modest increase to 5.72 ±0.13 × 10⁷ CFU.ml-1. As fermentation continued, S. thermophilus included a relatively stable population after 6 h, whereas L. bulgaricus grew, reaching a final count of 65.54 ±6.89 × 10⁷ CFU.ml-1 after 8 h. These findings were similar with those of Hansen et al., who verified synergistic growth of the two bacterial species through omics analyses [12]. This supported the idea that S. thermophilus initiated fermentation, whereas L. bulgaricus increased as fermentation advanced. Additionally, adaptability of these bacteria to various matrices was demonstrated by Nadeem et al., who showed that S. thermophilus and L. bulgaricus could effectively ferment plant-based milk alternatives. The current study highlighted changes in population sizes during fermentation and illustrated how the two bacteria acted together, with S. thermophilus starting the acidification process and creating conditions that allowing L. bulgaricus to increase, similar to previous studies.
- Conclusion
This study provides a broader understanding of fermentation in goat milks. It underscores the complem-entarity of S. thermophilus and L. bulgaricus. Generally, S. thermophilus contributes to the rapid acidification process, leading to the proliferation of L. bulgaricus, which is a significant contributor to the texture, stability and organ-oleptic characteristics of the final product. The synergistic interaction between these two species results in desirable texture and decreased syneresis. This improves quality of the fermented dairy products by enhancing the unique characteristics of goat milks. These findings are invaluable for optimizing production processes in the agricultural food industry, where the local context is critical particularly in use of natural resources, as exemplified by the use of Algerian goat milks. These advancements have further contributed to increasing demands for functional and healthy foods. Future studies should investigate interactions between these bacterial strains and other environmental parameters or natural ingredients. The major aim is to optimize nutritional and sensory qualities of the final products, while minimizing challenges such as excessive syneresis.
References
- Tayeb S, Saadia L, Hakim T, Abdellah F, Hamden K. Effect of milking frequency on the hygiene index and nutritional quality of raw milk: Mid-northern region Algerian study. Mod Phytol. 2025; 18: 9. https://doi.org/10.5281/zenodo.14619456
- Laws AP, Marshall VM. The relevance of exopolysaccharides to the rheological properties in milk fermented with ropy strains of lactic acid bacteria. Int Dairy J. 2001;11:709-721. https://doi.org/10.1016/S0958-6946(01)00115-7
- Canon F, Nidelet T, Guédon E, Thierry A, Gagnaire V. Understanding the Mechanisms of Positive Microbial Interactions That Benefit Lactic Acid Bacteria Co-cultures. Front Microbiol. 2020;11:1-16. https://doi.org/10.3389/fmicb.2020.02088
- Kekkonen RA, Lummela N, Karjalainen H, Latvala S, Tynkkynen S, Järvenpää S, et al. Probiotic intervention has strain-specific anti-inflammatory effects in healthy adults. World J Gastroenterol. 2008;14:2029-2036. https://doi.org/10.3748/wjg.14.2029
- Hols P, Hancy F, Fontaine L, Grossiord B, Prozzi D, Leblond-Bourget N, et al. New insights in the molecular biology and physiology of Streptococcus thermophilus revealed by comparative genomics. FEMS Microbiol Rev. 2005; 29:435-463. https://doi.org/10.1016/j.femsre.2005.04.008
- Crittenden RG, Martinez NR, Playne MJ. Synthesis and utilisation of folate by yoghurt starter cultures and probiotic bacteria. Int J Food Microbiol. 2003; 80: 217-222. https://doi.org/10.1016/S0168-1605(02)00170-8
- Qiu S, Zeng H, Yang Z, Hung W, Wang B, Yang A. Dynamic metagenome‐scale metabolic modeling of a yogurt bacterial community. Biotechnol Bioeng. 2023; 120: 2186-2198. https://doi.org/10.1002/BIT.28492
- International Organization for Standardization. ISO 6091:2010. Dried milk — Determination of titratable acidity (Reference method). 2nd ed. Geneva: ISO; 2010. Available from: https://www.iso.org/standard/55777.html
- International Organization for Standardization. ISO 554:2001. Standard atmospheres for conditioning and/or testing — Specifications. 1st ed. Geneva: ISO; 2001. Available from: https://www.iso.org/standard/29824.html
- International Organization for Standardization. ISO 17604:2015. Microbiology of the food chain — Carcass sampling for microbiological analysis. 2nd ed. Geneva: ISO; 2017. Available from: https://www.iso.org/standard/60313.html
- Moghadam SK, Farhoodi M, Mofid V, Homayouni-Rad A, Mortazavian AM, Bonab AM. Effects of Adding Non-viable Lacticaseibacillus casei and Lactobacillus acidophilus on Physicochemical, Microbial, Chemical and Sensory Attributes of Probiotic Doogh. Appl Food Biotechnol. 2024; 11: 1-15. https://doi.org/ 10.22037/afb.v11i1.44105
- Qiu S, Zeng H, Yang Z, Hung W-L, Wang B, Yang A. Dynamic community-level metabolic modeling for fermentation kinetics and metabolic interactions of the yogurt starter culture based on metagenomic analysis. Authorea (Authorea); 2023. https://doi.org/10.22541/au.167475604.47358393/v1
- Meng J, Liu J, Chen D, Li D, Wang J. Integration of metabolomics and transcriptomics reveals major metabolic pathways and potential biomarkers involved in aging mice with type 2 diabetes mellitus. Chin Med J (Engl). 2022; 135: 247-249. https://doi.org/10.1097/CM9.0000000000001554
- Ahsan S, Khaliq A, Chughtai MFJ, Nadeem M, Tahir A Bin, Din AA, et al. Technofunctional quality assessment of soymilk fermented with Lactobacillus acidophilus and Lactobacillus casei. Biotechnol Appl Biochem. 2022; 69: 172-182. https://doi.org/ 10.1002/BAB.2094
- Nadirova S, Sinyavskiy Y. Justification of the shelf life of dairy products based on goat’s milk. 2023; 49-57. https://doi.org/10.11134/btp.2.2023.4
- Sarvari F, Mortazavian AM, Fazeli MR. Biochemical characteristics and viability of probiotic and yogurt bacteria in yogurt during the fermentation and refrigerated storage. Appl Food Biotechnol. 2014; 1: 55-61. https://doi.org/ 10.22037/afb.v1i1.7125
- Sieuwerts S, Molenaar D, Van Hijum SAFT, Beerthuyzen M, Stevens MJA, Janssen PWM, et al. Mixed-Culture transcriptome analysis reveals the molecular basis of mixed-culture growth in Streptococcus thermophilus and Lactobacillus bulgaricus. Appl Environ Microbiol. 2010; 76: 7775-7784. https://doi.org/10.1128/AEM.01122-10
- Picon A, Garde S, Ávila M, Nuñez M. Microbiota dynamics and lactic acid bacteria biodiversity in raw goat milk cheeses. Int Dairy J. 2016;58:14-22. https://doi.org/10.1016/j.idairyj.2015.09.010
- Wu T, Guo S, Kwok LY, Zhang H, Wang J. Temperature-dependent metabolic interactions between Streptococcus thermophilus and Lactobacillus delbrueckii ssp. bulgaricus in milk fermentation: Insights from gas chromatography–ion mobility spectrometry metabolomics. J Dairy Sci. 2025; 108: 242-256. http://dx.doi.org/10.3168/jds.2024-25153
- Gaddour A, Najari S, Aroum S, Abdennebi M. Original Research Article Local goat milk valorization and dairy products characterization in the southern Tunisia arid zone. 2013; 2: 318-323.
- Sugata M, Kim Y, Rachmah AF, Oei J, Purnama ET, Nitbani ACJ, et al. Assessment of Probiotic Characteristics of Lactiplantibacillus plantarum SU-KC1a Isolated from Human Breast Milk in Indonesia. Appl Food Biotechnol. 2024; 11: 1-13. http://dx.doi.org/10.22037/afb.v11i1.45707
- Freire FC, Adorno MAT, Sakamoto IK, Antoniassi R, Chaves ACSD, dos Santos KMO, et al. Impact of multi-functional fermented goat milk beverage on gut microbiota in a dynamic colon model. Food Res Int. 2017; 99: 315-327. http://dx.doi.org/10.1016/J.FOODRES.2017.05.028
- Abbasalizadeh S, Hejazi MA, Hajiabbas MP. Kinetics of β-Galactosidase production by Lactobacillus bulgaricus during pH controlled batch fermentation in three commercial bulk starter media. Appl Food Biotechnol. 2015; 2: 39-47. http://dx.doi.org/10.22037/afb.v2i4.9512
- Sonnier JL, Karns JS, Lombard JE, Kopral CA, Haley BJ, Kim S-W, et al. Prevalence of Salmonella enterica, Listeria monocytogenes and pathogenic Escherichia coli in bulk tank milk and milk filters from US dairy operations in the National Animal Health Monitoring System Dairy 2014 study. J Dairy Sci. 2018; 101: 1943-1956. http://dx.doi.org/ 10.3168/jds.2017-13546
- Afzal A, Mahmood MS, Hussain I, Akhtar M. Adulteration and Microbiological Quality of Milk (A Review). Pakistan J Nutr. 2011; 10: 1195-1202. http://dx.doi.org/10.3923/pjn.2011.1195.1202
- Runge MO, Bloch S, Han H. Composition and process for producing a fermented milk product comprising application of a lactose-deficient S. thermophilus strain, a lactose-deficient L. bulgaricus strain and a probiotic strain. US Patent US20210161163A1. Published 2021 Jun 3. Available from: https://patentimages.storage.googleapis.com/a4/fe/ae/9ff192ccb431c6/US20210161163A1.pdf
- Acidification
- Fermentation
- Goat milk
- Lactobacillus bulgaricus
- Streptococcus thermophilus
- Viscosity
##submission.howToCite##
参考
References
Tayeb S, Saadia L, Hakim T, Abdellah F, Hamden K. Effect of milking frequency on the hygiene index and nutritional quality of raw milk: Mid-northern region Algerian study. Mod Phytol. 2025; 18: 9. https://doi.org/10.5281/zenodo.14619456
Laws AP, Marshall VM. The relevance of exopolysaccharides to the rheological properties in milk fermented with ropy strains of lactic acid bacteria. Int Dairy J. 2001;11:709-721. https://doi.org/10.1016/S0958-6946(01)00115-7
Canon F, Nidelet T, Guédon E, Thierry A, Gagnaire V. Understanding the Mechanisms of Positive Microbial Interactions That Benefit Lactic Acid Bacteria Co-cultures. Front Microbiol. 2020;11:1-16. https://doi.org/10.3389/fmicb.2020.02088
Kekkonen RA, Lummela N, Karjalainen H, Latvala S, Tynkkynen S, Järvenpää S, et al. Probiotic intervention has strain-specific anti-inflammatory effects in healthy adults. World J Gastroenterol. 2008;14:2029-2036. https://doi.org/10.3748/wjg.14.2029
Hols P, Hancy F, Fontaine L, Grossiord B, Prozzi D, Leblond-Bourget N, et al. New insights in the molecular biology and physiology of Streptococcus thermophilus revealed by comparative genomics. FEMS Microbiol Rev. 2005; 29:435-463. https://doi.org/10.1016/j.femsre.2005.04.008
Crittenden RG, Martinez NR, Playne MJ. Synthesis and utilisation of folate by yoghurt starter cultures and probiotic bacteria. Int J Food Microbiol. 2003; 80: 217-222. https://doi.org/10.1016/S0168-1605(02)00170-8
Qiu S, Zeng H, Yang Z, Hung W, Wang B, Yang A. Dynamic metagenome‐scale metabolic modeling of a yogurt bacterial community. Biotechnol Bioeng. 2023; 120: 2186-2198. https://doi.org/10.1002/BIT.28492
International Organization for Standardization. ISO 6091:2010. Dried milk — Determination of titratable acidity (Reference method). 2nd ed. Geneva: ISO; 2010. Available from: https://www.iso.org/standard/55777.html
International Organization for Standardization. ISO 554:2001. Standard atmospheres for conditioning and/or testing — Specifications. 1st ed. Geneva: ISO; 2001. Available from: https://www.iso.org/standard/29824.html
International Organization for Standardization. ISO 17604:2015. Microbiology of the food chain — Carcass sampling for microbiological analysis. 2nd ed. Geneva: ISO; 2017. Available from: https://www.iso.org/standard/60313.html
Moghadam SK, Farhoodi M, Mofid V, Homayouni-Rad A, Mortazavian AM, Bonab AM. Effects of Adding Non-viable Lacticaseibacillus casei and Lactobacillus acidophilus on Physicochemical, Microbial, Chemical and Sensory Attributes of Probiotic Doogh. Appl Food Biotechnol. 2024; 11: 1-15. https://doi.org/ 10.22037/afb.v11i1.44105
Qiu S, Zeng H, Yang Z, Hung W-L, Wang B, Yang A. Dynamic community-level metabolic modeling for fermentation kinetics and metabolic interactions of the yogurt starter culture based on metagenomic analysis. Authorea (Authorea); 2023. https://doi.org/10.22541/au.167475604.47358393/v1
Meng J, Liu J, Chen D, Li D, Wang J. Integration of metabolomics and transcriptomics reveals major metabolic pathways and potential biomarkers involved in aging mice with type 2 diabetes mellitus. Chin Med J (Engl). 2022; 135: 247-249. https://doi.org/10.1097/CM9.0000000000001554
Ahsan S, Khaliq A, Chughtai MFJ, Nadeem M, Tahir A Bin, Din AA, et al. Technofunctional quality assessment of soymilk fermented with Lactobacillus acidophilus and Lactobacillus casei. Biotechnol Appl Biochem. 2022; 69: 172-182. https://doi.org/ 10.1002/BAB.2094
Nadirova S, Sinyavskiy Y. Justification of the shelf life of dairy products based on goat’s milk. 2023; 49-57. https://doi.org/10.11134/btp.2.2023.4
Sarvari F, Mortazavian AM, Fazeli MR. Biochemical characteristics and viability of probiotic and yogurt bacteria in yogurt during the fermentation and refrigerated storage. Appl Food Biotechnol. 2014; 1: 55-61. https://doi.org/ 10.22037/afb.v1i1.7125
Sieuwerts S, Molenaar D, Van Hijum SAFT, Beerthuyzen M, Stevens MJA, Janssen PWM, et al. Mixed-Culture transcriptome analysis reveals the molecular basis of mixed-culture growth in Streptococcus thermophilus and Lactobacillus bulgaricus. Appl Environ Microbiol. 2010; 76: 7775-7784. https://doi.org/10.1128/AEM.01122-10
Picon A, Garde S, Ávila M, Nuñez M. Microbiota dynamics and lactic acid bacteria biodiversity in raw goat milk cheeses. Int Dairy J. 2016;58:14-22. https://doi.org/10.1016/j.idairyj.2015.09.010
Wu T, Guo S, Kwok LY, Zhang H, Wang J. Temperature-dependent metabolic interactions between Streptococcus thermophilus and Lactobacillus delbrueckii ssp. bulgaricus in milk fermentation: Insights from gas chromatography–ion mobility spectrometry metabolomics. J Dairy Sci. 2025; 108: 242-256. http://dx.doi.org/10.3168/jds.2024-25153
Gaddour A, Najari S, Aroum S, Abdennebi M. Original Research Article Local goat milk valorization and dairy products characterization in the southern Tunisia arid zone. 2013; 2: 318-323.
Sugata M, Kim Y, Rachmah AF, Oei J, Purnama ET, Nitbani ACJ, et al. Assessment of Probiotic Characteristics of Lactiplantibacillus plantarum SU-KC1a Isolated from Human Breast Milk in Indonesia. Appl Food Biotechnol. 2024; 11: 1-13. http://dx.doi.org/10.22037/afb.v11i1.45707
Freire FC, Adorno MAT, Sakamoto IK, Antoniassi R, Chaves ACSD, dos Santos KMO, et al. Impact of multi-functional fermented goat milk beverage on gut microbiota in a dynamic colon model. Food Res Int. 2017; 99: 315-327. http://dx.doi.org/10.1016/J.FOODRES.2017.05.028
Abbasalizadeh S, Hejazi MA, Hajiabbas MP. Kinetics of β-Galactosidase production by Lactobacillus bulgaricus during pH controlled batch fermentation in three commercial bulk starter media. Appl Food Biotechnol. 2015; 2: 39-47. http://dx.doi.org/10.22037/afb.v2i4.9512
Sonnier JL, Karns JS, Lombard JE, Kopral CA, Haley BJ, Kim S-W, et al. Prevalence of Salmonella enterica, Listeria monocytogenes and pathogenic Escherichia coli in bulk tank milk and milk filters from US dairy operations in the National Animal Health Monitoring System Dairy 2014 study. J Dairy Sci. 2018; 101: 1943-1956. http://dx.doi.org/ 10.3168/jds.2017-13546
Afzal A, Mahmood MS, Hussain I, Akhtar M. Adulteration and Microbiological Quality of Milk (A Review). Pakistan J Nutr. 2011; 10: 1195-1202. http://dx.doi.org/10.3923/pjn.2011.1195.1202
Runge MO, Bloch S, Han H. Composition and process for producing a fermented milk product comprising application of a lactose-deficient S. thermophilus strain, a lactose-deficient L. bulgaricus strain and a probiotic strain. US Patent US20210161163A1. Published 2021 Jun 3. Available from: https://patentimages.storage.googleapis.com/a4/fe/ae/9ff192ccb431c6/US20210161163A1.pdf
- 摘要 ##plugins.themes.ojsPlusA.frontend.article.viewed##: 574 ##plugins.themes.ojsPlusA.frontend.article.times##
- pdf (English) ##plugins.themes.ojsPlusA.frontend.article.downloaded##: 267 ##plugins.themes.ojsPlusA.frontend.article.times##