
Design of Statistically-Based Bioprocesses for the Enhanced Production of Moderate Thermophilic Alkaline Α-Amylase from Bacillus Subtilis Isolated From Guilan Rice Mill Wastes
بیوتکنولوژی غذایی کاربردی,
دوره 12 شماره 1 (2025),
4 دی 2025
,
صفحه 1-11 (e26)
https://doi.org/10.22037/afb.v12i1.50183
چکیده
Background and Objective: The demand for cost-effective and thermostable α-amylases for industrial applications has driven the research to discover new microbial sources. This research aimed to isolate and characterize α-amylase-producing bacteria from rice milling wastes and employ Response Surface Methodology (RSM) to improve enzyme production.
Material and Methods: Bacterial samples were collected from different agro-industrial wastes and primarily screened using Lugol's iodine method. Secondary isolation was performed by α-amylase activity assessment using the DNS assay. Enzyme production was optimized by RSM, with the temperature, pH, and starch concentration as key variables. In addition, the effect of different pH and temperatures was assessed on the α-amylase activity. 16S rRNA sequencing and phylogenetic analysis were used for bacterial identification.
Results and Conclusion: The isolate was identified as Bacillus subtilis NllST B 627. Optimum conditions for maximum enzyme production (0.21 Umg-1) were starch 5.5 gL-1, temperature 40°C, and pH 7. Temperature was the most significant factor influencing enzyme production, whereas pH and starch concentration showed weaker effects but potentially relevant interactions. The overall model based on response surface curves was statistically significant, indicating that the combination of independent variables significantly influences enzyme production. The enzyme exhibited maximum activity at pH 7, while the lowest activity was observed at pH 5. Also, the enzyme's optimal activity occurred at 40°C, while the lowest catalysis was detected at 60°C. The identified strain exhibits promising properties for application in starch hydrolysis and other industrial purposes. This highlights the potential of rice mill wastes as a sustainable and low-cost resource for microbial enzyme production, and this study is the first to explore Guilan rice mill wastes for α-amylase production.
Keywords: α-Αmylase production, Detergent industry, Enzyme optimization, Isolation and identification, RSM Method
Introduction
Producing enzymes on an industrial scale is an expensive effort. A potential approach to overcome this issue involves identifying enzyme-producing microorganisms from agricultural/industrial waste, a method that both lowers production costs and promotes waste beneficial reuse and environmental sustainability [1,2]. So, rice milling wastes are an abundant by-product found in areas where rice is cultivated and constitute a largely available material full of microbial diversity. The moist and starch-rich remnants produced during rice processing create a perfect environment for the growth of α-amylase-producing bacteria, which have potential uses in various industries [3].
α-Αmylases (EC 3.2.1.1) are extensively utilized in the food, fermentation, detergent, pharmaceutical industries, ethanol production [4], and as an antibiofilm agent [5]. These enzymes cleave α-1,4-glycosidic bonds in starch, facilitating the production of various products such as dextrose, glucose, and starch syrups [6]. Although many thermostable α-amylase-producing strains, such as Bacillus subtilis, Bacillus amyloliquefaciens, Bacillus stearothermo-philus, Bacillus licheniformis, Bacillus polymyxa, and Bacillus coagulans have been identified so far [7], discovering new strains from novel sources could result in enzymes operating in extreme conditions. This leads to increasing efficiency and reducing costs on an industrial scale [8].
Globally, α-amylases account for approximately 23-33% of the enzymatic market share [9]. However, the cost of industrial-scale enzyme production remains significantly high. It is estimated that microbial culture media formulation alone accounts for about 30-40% of the final enzyme production costs [10]. Consequently, there is a persistent need to reduce these costs by developing inexpensive culture media formulations, leading to significant efforts to identify cost-effective alternatives for industrial enzyme production.
Various studies have been conducted on the identification and isolation of α-amylase-producing strains from different sources, including agricultural soils such as potato fields [11], industrial soils such as brick kiln [12], hot spring [13], deserts [14], different wastes [15,16], etc. For example, Niyomukiza et al. isolated amylolytic bacteria from starchy food wastes (maize meal and potato peel wastes), and 16S rRNA sequencing verified them as Bacillus subtilis. The optimum temperature for the enzyme was 60 °C, and the pH was 9 [17]. Tripathi et al. used Bacillus polymyxa NCIM 2539 to produce amylase using agro-industrial byproducts. Among various substrates tested, orange peel yielded the highest enzyme activity. Supplementation of the medium significantly enhanced amylase production, with optimal levels obtained at specific concentrations of orange peel, cysteine, and thiamine [18].
Many industries produce agricultural waste, one of which is the rice factory industry located in the North of Iran. They produce waste with high organic matter content, one of which is starch. The starch contained in the liquid waste can be broken down by bacteria-producing amylase enzymes into simpler molecules. In this research, there has been scarce focus on the microbial strains that exist within the starch-rich waste produced by rice mills, especially in Guilan province (Iran), where rice farming is a significant agricultural profession. This research aimed to isolate and molecularly characterize α-amylase-producing bacteria from rice milling waste and employ Response Surface Methodology (RSM) to improve enzyme production processes. Optimization of three parameters (incubation time, starch concentration, and incubation temperature) was investigated. Using an inexpensive, starch-rich waste substrate and statistical modeling to enhance enzyme output, this study could contribute to cost-effective enzyme development and play a significant role in sustainable industrial waste management.
- Materials and Methods
2.1. Primary isolation and screening of α-amylase-producing bacteria
Bacterial samples were collected from rice mill wastes, chip manufacturing wastes, bakery dusts, textile wastewater, and other industrial waste, because these sources contain a significant percentage of starch and are likely to contain more degrading bacteria. For isolating spore-forming bacteria, particularly Bacillus species, serial dilutions were prepared. Heat shock treatment was applied for 10 minutes at 80°C in a water bath. Subsequently, bacteria were isolated using the streak plate method on Nutrient Agar medium [19]. The isolated bacteria were then tested for starch hydrolysis by inoculating them onto the starch agar plates containing 2% starch and incubating for 24 hours. After incubation, the ability to hydrolyze starch was assessed by flooding the plates with Lugol's iodine solution. Bacteria showing a clear zone around colonies, indicative of starch hydrolysis, were selected for further analysis.
2.2. Secondary screening by α-amylase activity assessment
In the secondary screening phase, bacterial isolates that produced larger clear zones were selected for further enzymatic activity assessment. The α-amylase activity was measured in 250 mL flasks containing 50 mL of cultivation medium with the following composition (gL-1, Sigma-Aldrich, USA): starch 10%, peptone 5%, yeast extract 2.05%, NaCl 1.5 grL-1, KH₂PO₄ 0.5 gr/L, MgSO₄ 0.5 grL-1, CaCl₂ 0.1 grL-1, and glycerol 15% (vv-1). The flasks were incubated on a shaker at 120 rpm and 37°C for 48 hours. The initial pH of the medium was adjusted to 7.0. The inoculated bacterial strains were transferred into sterile pre-prepared medium. After incubation, samples were centrifuged at 10,000 rpm for 20 minutes, and the supernatant was collected for enzyme activity measurement.
2.3. Enzyme activity determination using the DNS method
The dinitrosalicylic Acid (DNS) method was employed to quantify α-amylase activity [20]. DNS is an alkaline reagent that reacts with reducing sugars, causing a color change from yellow to reddish-brown. A reaction mixture comprising 0.5 mL of crude enzyme and 0.5 mL of 1% starch solution was prepared. The mixture was incubated at 37°C for 30 minutes. The reaction was stopped by adding 1 mL of DNS reagent, followed by boiling for 5 minutes. Glucose concentration was measured using a spectrophoto-meter at 540 nm. A glucose standard curve was generated by plotting absorbance at 540 nm against the amounts of glucose released to define the concentration of glucose formed in each solution. One unit of enzyme activity was defined as the amount of enzyme required to liberate 1 μmol of reducing sugars per minute.
2.4. Response surface methodology for production optimization
The three strains exhibiting the highest production levels were selected from the screened bacterial isolates, and strain 2 was employed for statistical production optimization. RSM is a technique used to evaluate the influence of input parameters on responses [21]. Central Composite Design (CCD) provides an efficient approach to predict the interaction effects of influential factors on the process. In this study, optimization aimed to maximize enzyme production, using the Statistical Design-Expert 7.0 software, followed by CCD analysis [22].
In this design, the quantitative impact of the most effective variables, including starch concentration, temperature, and pH, were examined (Table 1). All experiments were conducted in triplicate. Positive and negative control strains were processed with every batch.
2.5. Evaluation of pH on the α-amylase activity
The bacteria were cultivated in production media and incubated in a shaker incubator for 24 hours, followed by centrifugation. In separate tubes, 0.5 mL of the supernatant was mixed with 0.5 mL of sodium carbonate buffers at 50 mM with pH values of 9 and 10, trisodium citrate buffers at 50 mM with pH 5 and 6, disodium hydrogen phosphate buffers at 50 mM with pH 7 and 8, and starch. The mixtures were incubated at 37°C for 30 minutes. Subsequently, 0.5 mL of DNS reagent was added, and absorbance was measured at 540 nm using a spectrophotometer.
2.6. Evaluation of temperature on the α-amylase activity
To evaluate the effect of temperature, bacteria were cultured in the production medium, centrifuged, and then 0.5 mL of the supernatant was mixed with 0.5 mL of 1% starch solution in a test tube. The test tubes were then placed in a water bath or thermostatic water bath set at temperatures of 40°, 50°, 60°, and 70°C for 30 minutes. The enzyme activity under each condition was subsequently assessed.
- Identification of α-amylase-producing bacteria
α-Amylase-producing bacteria were identified through phenotypic and macroscopic characterization, followed by microscopic examination using Gram staining and sporulation tests. For definitive identification, 16S rRNA gene analysis was conducted. Bacterial DNA was extracted using a boiling method. Briefly, the bacterial cultures were centrifuged at 2000 g for 20 minutes to obtain a cell pellet. The pellet was resuspended in sterile distilled water, and the microtubes were first placed in a freezer and then boiled in a water bath to lyse the bacterial cells and release the DNA. After centrifugation at 7000 rpm for 10 minutes, the supernatant containing the DNA was collected and precipitated with cold ethanol. The DNA pellet was air-dried and dissolved in a small volume of distilled water, and its concentration and purity were measured using a NanoDrop spectrophotometer. PCR amplification was performed with universal bacterial primers 27F (5′- AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-CGGTTACCTTGTTACGACTT-3′) [23]. The PCR protocol consisted of 30 cycles involving: denaturation at 95°C for 1 minute, annealing at 56°C for 1 minute, extension at 72°C for 1 minute, and a final elongation step at 72°C for 7 minutes. The PCR products were analyzed by gel electrophoresis, and successful amplicons were sent to Pishgaman Co. (Iran, Tehran) for sequencing. The obtained sequences were compared to those in the NCBI database using the BLAST tool (https://blast.ncbi.nlm.nih-gov/Blast.cgi) [24]. The closest related species were identified based on the similarities in the 16S rRNA gene sequences of the α-amylase. For phylogenetic analysis, the sequences were searched and aligned in the NCBI GenBank database using multiple sequence alignment with the ClustalW algorithm (https://www.genome.jp/tools-bin/clustalw). A phylogeny-etic tree was constructed using the Molecular Evolution Genetic Analysis (MEGA 4) software with the Neighbor-Joining algorithm and 1000 bootstrap replicates [25].
- Statistical analysis
Data were analyzed using the Design Expert 7.0 software. All the experiments were conducted in triplicate. Results are presented as mean values ± standard error (SE), with a significance threshold of P < 0.05.
- Results and Discussion
3.1. Isolation and screening of α-amylase-producing bacteria
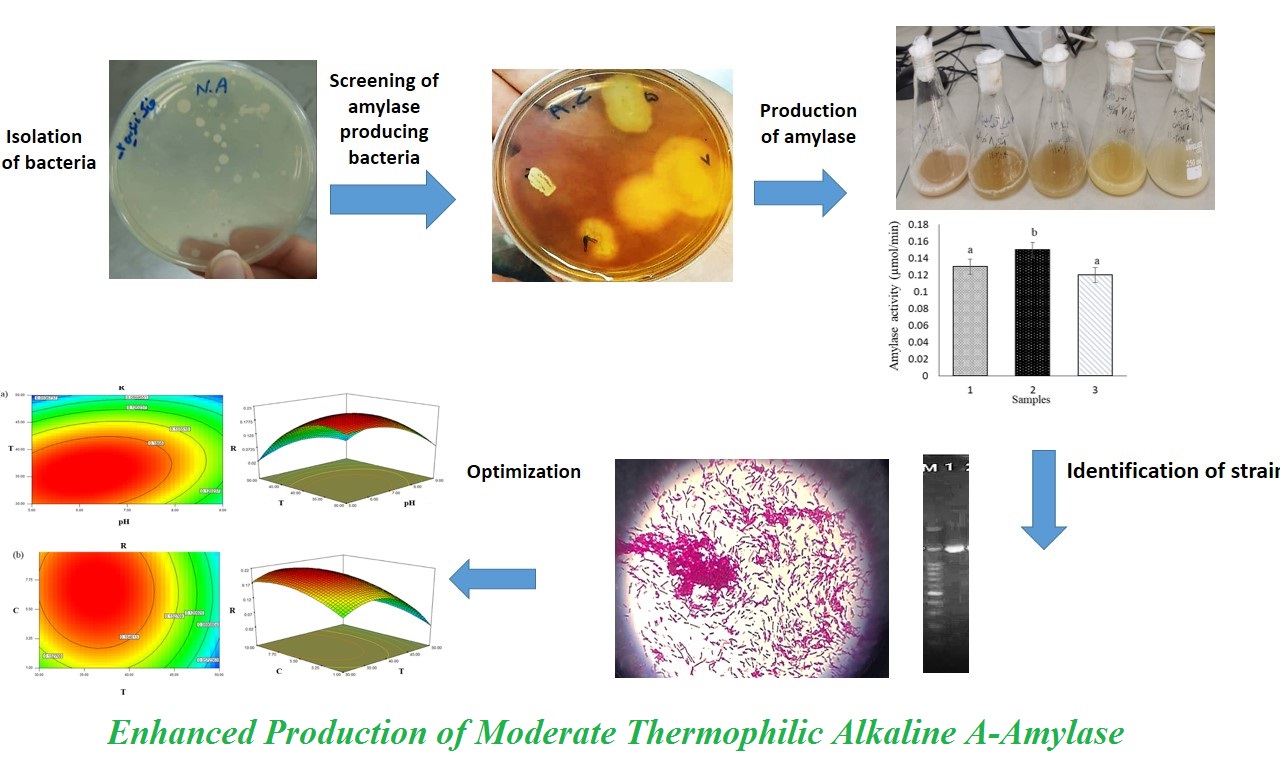
The Lugol's iodine test was performed for primary isolation of α-amylase-producing bacteria from different waste samples. Results demonstrated that three bacterial isolates, including strain #1 from the bakery's waste, strain #2 from the rice mill's waste, and strain #3 from the agricultural waste, produced extracellular α-amylase, positively (Figure 1 A to C), as evidenced by the presence of a clear zone surrounding the colonies after washing with iodine solution. The formation of a clear zone indicates that the starch present in the starch agar medium was hydrolyzed into monomers by α-amylase enzymes produced by the bacteria. In contrast, a dark blue-black zone represents an iodine-starch complex. The isolates showing clear zones were considered positive for α-amylase production (Figure 1 A to C). In contrast, the absence of a clear zone indicated a lack of extracellular α-amylase activity (Figure 1D).
3.2. Assessment of bacterial α-amylase activity
α-Amylase activity of the three selected samples was assessed for further screening of bacteria. Results revealed that the α-amylase activity of strain #2, from the rice mill waste, was significantly higher than that of others (P < 0.05) (Figure 2).
3.3. Experimental design
In this design, based on RSM and using the CCD method, 15 experimental runs were arranged, as summarized in Table 2. In this research, based on a multiple regression analysis of experimental data, the final model was presented as follows (Equation 1):
α-Amylase activity = 0.20 - 0.025A - 0.055B +0.020C + 0.030AB - 0.025AC - -5.000E-003BC - 0.046A2 - 0.076B2 - 0.031C2
Where A, B, and C are pH, starch concentration, and temperature, respectively.
The analysis of variance (ANOVA) is described in Tables 3 and 4. The larger the F-value, the greater the variation between sample means relative to the variation within the samples. The p-value is the probability of obtaining an F-ratio as large or larger than the one observed, assuming that there is no difference between the group averages. The model F value of 51.62 implies that the model is significant. Values of Prob > F less than 0.05 imply that the model terms are relevant at the 95% confidence level. The model showed a high determination coefficient (R2 = 0.9894), indicating a strong correlation between the experimental and predicted values.
Figure 3 shows the normal plots of residual (difference between the observed and predicted value). A low residual value is necessary for a good mathematical model fitted on observed data. The predicted responses and observed responses are shown in Figure 4 and the data points located close to the diagonal line, suggesting a satisfactory correlation.
To study the interaction among the different independent variables and their corresponding effect on the response, contour plots and 3-D plots were drawn. A contour plot is a graphical representation of a three-dimensional response surface based on two independent variables, helping to illustrate their main and interaction effects on the response. Figure 5 shows the amylase response and correlation between variables in plots. Optimum conditions for maximum enzyme production (0.21 Umg-1) were starch 5.5 gL-1, temperature 40°C, and pH 7.
The bell-shaped surface indicates that extreme acidic or alkaline conditions and temperatures above 45 °C negatively influence production, likely due to reduced microbial growth or enzyme instability. The nearly symmetrical surface curvature also suggests that the process remains relatively stable near the optimal point.
In Fig. 5b, the interaction between temperature and starch concentration shows that enzyme yield rose with increasing starch up to about 5–6 gL-1, but declined at higher levels, possibly because of substrate inhibition or catabolized repression. Maximum activity was achieved at a moderate starch concentration and 40 °C. Together, these plots confirm that temperature is the dominant factor, while pH and substrate concentration contribute secondary but interactive effects, defining a narrow yet stable region for optimal amylase production. According to the results of 15 experiments, the lowest activity was observed at high temperature, low starch concentration, and under alkaline pH. This suggests excessive heat or inappropriate substrate levels may negatively impact enzyme production. These data underline that interactions between parameters are critical in optimizing enzyme yield, and single-factor optimization may be insufficient. Temperature was identified as the most significant factor influencing enzyme production, whereas pH and starch concentration showed weaker individual effects but potentially relevant interactions. When compared with similar studies, some differences in optimal conditions were observed. In the work performed by Adetiloye et al., Bacillus cereus from a warm spring demonstrated a slightly higher optimal temperature at 45°C and an RSM-predicted optimal pH of 7, although OFAT analysis also indicated potent activity at pH 8 [23]. In the study performed by Sharif et al., Bacillus licheniformis exhibited an even higher optimal temperature at 55°C and a more alkaline preference at pH 9. This suggests that variations in enzyme thermostability and pH tolerance may depend on the species [26].
Regarding substrate concentration, both the current study and the work by Adetiloye et al. [23] identified 5% starch as optimal, whereas Sharif et al. reported an optimum of 1% [26], possibly due to substrate inhibition effects at higher concentrations. The differences in assay methods (µmolmin-1 vs. UmL-1) and strain-specific enzyme kinetics make direct comparison of activity values difficult; however, both Adetiloye et al. and Sharif et al. reported higher numerical activities than the current study, which could be related to strain genetics, cultivation conditions, or methodological variations in activity measurement. The emphasis is on identifying robust operating conditions within the region of interest that maximize production while remaining practically feasible in downstream processing.
3.4. The effect of pH on the α-amylase activity
As illustrated in Figure 6A, the enzyme exhibited maximum activity at pH 8, while the lowest activity was observed at pH 5 (P < 0.05). It can be concluded that the enzyme operates most efficiently under slightly alkaline conditions, which may be due to the stabilization of the enzyme’s active site and overall tertiary structure at this pH, which enhances its catalytic efficiency. Conversely, the lowest enzymatic activity at pH 5 suggests that acidic conditions lead to reduced enzyme performance, likely as a result of denaturation or alteration in the ionization state of critical amino acids involved in substrate binding and catalysis [27]. A moderate reduction in the activity at pH 9 indicates a narrow optimal pH range for the enzyme. These findings emphasize the importance of pH optimization in industrial applications of this enzyme. These findings are consistent with previously reported characteristics of bacterial α-amylases, which generally prefer neutral to slightly alkaline environments for optimal activity [14,22,28,29].
3.5. The effect of temperature on the α-amylase activity
The impact of temperature on the catalytic activity of α-amylase from Isolate 2 is presented in Figure 6B. The enzyme demonstrated optimal activity at 50°C, while the lowest enzyme activity was detected at 60°C (P < 0.05). The enzyme may retain its functional conformation at moderately high temperatures. This property makes the enzyme potentially suitable for industrial processes that require elevated temperatures, such as starch liquefaction and food processing [30]. Reduction of catalytic activity at 60°C may be due to thermal denaturation or irreversible structural changes, leading to loss of function [27]. Overall, the results confirm that temperature plays a critical role in the amylolysis function, and identifying the optimum point is crucial for maximizing enzymatic yield.
Various studies have reported varying optimal temperatures and pHs for the α-amylase activity of Bacillus species isolated from different sources. Some of them are as follows: The highest α-amylase activity of the purified Bacillus licheniformis strain LB04 isolated from Espinazo hot springs in Mexico was at pH 3 and 65 ºC [31]. Bacillus licheniformis HULUB1 and Bacillus subtilis SUNGB2 isolated from Malaysian hot spring showed the highest α-amylase activity at 65° C and pH 6.0 [13]. Bacillus cereus and Bacillus licheniformis isolated from the potato fields demonstrated α-amylase function at pH 8.0 and temperatures of 45°C and 65°C, respectively [11]. Maximum α-amylase activity of the three isolates of Bacillus from a hot place in Ethiopia occurred at 75°C, 70°C, and 65°C, and the highest activity was at pH 8 [14]. The highest α-amylase activity of B. subtilis and B. licheniformis isolated from different soil samples in Punjab (Pakistan) was recorded at 70°C and pH 9 [22].
3.6. Molecular identification and phylogenetic analysis of the strain
The 16S rRNA gene was employed for the bacterial isolate molecular identification. Genomic DNA was extracted and subsequently used as a template for PCR amplification. The amplified product was then subjected to DNA sequencing, and the resulting sequence was analyzed using the BLASTn tool (NCBI) to compare it with known bacterial sequences. The isolate showed 100% sequence similarity with Bacillus subtilis, indicating that the strain belongs to this species. More specifically, the sequence demonstrated the highest similarity to Bacillus subtilis strain NllST B 627, with no nucleotide mismatches observed, further confirming the identity of the isolate.
To validate the BLAST results and investigate the strain evolutionary relationship, a phylogenetic tree was constructed using the neighbor-joining method based on 16S rRNA sequences of closely related strains (Figure 7). The tree showed that the isolate clustered tightly with Bacillus subtilis strains, particularly with strain NllST B 627, further supporting the identification. The accurate identification of the strain is crucial for further biotechnological applications and strain improvement strategies.
Although this study successfully identified and optimized α-amylase production from Bacillus subtilis, several limitations should be mentioned. Enzyme purification was not performed by chromatographic methods. The thermal and pH stability of the enzyme were not assessed, which are critical parameters for its industrial applicability. No direct comparison was made with commercially available or industrial reference strains. The effects of metal ions, inhibitors, and activators on enzyme activity were not evaluated, which could provide valuable insight into enzyme regulation and potential process enhancements. Future studies should focus on a comprehensive characterization of the enzyme, including stability profiling, kinetic analysis, and evaluation under various physicochemical and chemical conditions.
- Conclusion
This study successfully isolated and characterized a moderate thermostable alkaline α-amylase-producing Bacillus subtilis strain from rice mill waste and optimized its enzyme production using RSM. Maximum enzyme production (0.21 Umg-1) was acheived with starch 5.5 gL-1, temperature 40°C, and pH 7. The enzyme displayed optimal activity at a pH of 8 and 50°C. Temperature exerted the most significant effect on enzyme production. These findings emphasize the possibility of employing agricultural by-products such as rice mill waste for low-cost microbial enzyme production while contributing to sustainable waste management. Although further purification, stability testing, and scale-up studies are needed, the identified strain represents a promising candidate for applications in the detergent, food, and biotechnological industries. Coupled isolation from an environmental waste source with statistical optimization (RSM/CCD) to achieve a robust production process, illustrating a streamlined path from waste to biocatalyst. Evaluation of production and recovery in pilot-scale bioreactors, including downstream processing feasibility and cost analysis will be suggested.
- Acknowledgements
We thank the research core of Yazd University for the financial support during the course of this project.
- Declaration of competing interest
The authors declare no conflict of interests.
- Authors’ Contributions
Conceptualization: Mirbagheri and Chamani; investigation: Alkoozei, methodology: Mirbagheri and Khatami; Data curation: Mirbagheri; writing: Chamani and Mirbagheri; review and editing: Khatami; Supervision: Mirbagheri and Chamani.
- Using Artificial Intelligent Chatbots
The authors declare that Artificial Intelligence (AI)-assisted tools were used solely for language editing, grammar checking, and improving readability of the manuscript. All scientific content and writing were generated by the authors themselves, who take full responsibility for the integrity and accuracy of the work.
- Ethical Consideration
This study focused on bacteria and did not involve any human or animal subjects; no human data or samples were used.
- α-Αmylase production
- Detergent industry
- Enzyme optimization
- Isolation and identification
- RSM Method
ارجاع به مقاله
مراجع
Sanjaya EH, Suharti S, Alvionita M, Telussa I, Febriana S, Clevanota H. Isolation and characterization of amylase enzyme produced by indigenous bacteria from sugar factory waste. Open Biotechnol J. 2024; 18. https://doi.org/10.2174/0118740707296261240418114958
Masi C, Tebiso A, Selva Kumar K V. Isolation and characterization of potential multiple extracellular enzyme-producing bacteria from waste dumping area in Addis Ababa. Heliyon. 2023; 9. https://doi.org/10.1016/j.heliyon.2022.e12645
Sharma V, Tsai M-L, Nargotra P, Chen C-W, Kuo C-H, Sun P-P, et al. Agro-industrial food waste as a low-cost substrate for sustainable production of industrial enzymes: A critical review. Catalysts. 2022; 12. https://doi.org/10.3390/catal12111373
Balcerek M. Polysaccharides of starchy and lignocellulose materials and their use in ethanol production: Enzymes and other factors affecting the production process. Appl Food Biotechnol. 2022; 9: 157-172. https://doi.org/10.22037/afb.v9i2.37355
Al Ghamdi H, Zabermawi N, Aly MM. Biofilm formation of foodborne pathogens and strategies of its prevention and biocontrol: A review. Appl Food Biotechnol. 2024; 12: 1-8. https://doi.org/10.22037/afb.v12i1.46861
Ashok PP, Dasgupta D, Ray A, Suman SK. Challenges and prospects of microbial α-amylases for industrial application: a review. World J Microbiol Biotechnol. 2024; 40: 44. https://doi.org/10.1007/s11274-023-03821-y
Kikani BA, Singh SP. Amylases from thermophilic bacteria: structure and function relationship. Crit Rev Biotechnol. 2022; 42: 325-341. https://doi.org/10.1080/07388551.2021.1940089
Jaiswal N, Jaiswal P. Thermostable α-amylases and laccases: Paving the way for sustainable industrial applications. Processes 2024; 12. https://doi.org/10.3390/pr12071341
Paul JS, Gupta N, Beliya E, Tiwari S, Jadhav SK. Aspects and recent trends in microbial α-amylase: A review. Appl Biochem Biotechnol. 2021; 193: 2649-2698. https://doi.org/10.1007/s12010-021-03546-4
Nurfarahin AH, Mohamed MS, Phang LY. Culture medium development for microbial-derived surfactants production—An overview. Molecules. 2018; 23: 1049. https://doi.org/10.3390/molecules23051049
Rakaz MA, Hussien MO, Ibrahim HM. Isolation, extraction, purification, and molecular characterization for thermostable α -amylase from locally isolated bacillus species in sudan. Biochem Res Int. 2021; 2021: 6670380. https://doi.org/10.1155/2021/6670380
Ullah I, Khan MS, Khan SS, Ahmad W, Zheng L, Shah SUA, et al. Identification and characterization of thermophilic amylase producing bacterial isolates from the brick kiln soil. Saudi J Biol Sci. 2021; 28 :970-979. https://doi.org/10.1016/j.sjbs.2020.11.017
Msarah MJ, Ibrahim I, Hamid AA, Aqma WS. Optimisation and production of alpha amylase from thermophilic Bacillus spp. and its application in food waste biodegradation. Heliyon. 2020; 6. https://doi.org/10.1016/j.heliyon.2020.e04183
Yassin SN, Jiru TM, Indracanti M. Screening and characterization of thermostable amylase‐producing bacteria isolated from soil samples of afdera, afar region, and molecular detection of amylase‐coding gene. Int J Microbiol. 2021; 2021: 5592885. https://doi.org/10.1155/2021/5592885
Okunwaye T, Uadia PO, Okogbenin BO, Okogbenin EA, Onyia DC, Obibuzor JU. Amylase-producing fungi and bacteria associated with some food processing wastes. Niger J Biotechnol. 2021; 38: 74-82. https://doi.org/10.4314/njb.v38i1.9
Isabel JB, Premalatha M. Harnessing amylase producing novel bacterial strains from industrial waste: A bioprocessing and optimization approach. Int J Biol Macromol. 2025: 146769. https://doi.org/10.1016/j.ijbiomac.2025.146769
Niyomukiza S, Owino W, Mathara JM, Maina N. Isolation, purification and biochemical characterization of alkaline α-amylase from Bacillus subtilis strain W3SFR5 isolated from kitchen wastes. Appl Food Biotechnol. 2022; 10:9-19. https://doi.org/10.22037/afb.v10i1.39495
Tripathi AD, Joshi A, Singh SP, Shrivastava A. Production of amylase by bacillus polymyxa ncim no. 2539 from agroindustrial wastes. Appl Food Biotechnol. 2017; 4: 103-112. https://doi.org/10.22037/afb.v4i2.15471
Akhbariyoon HR, Mirbagheri M, Emtiazi G. Isolation and identification of Alicyclobacillus with high dipicolinic acid and heat resistant proteins from mango juice. Appl Food Biotechnol. 2016; 3: 270-274 https://doi.org/10.22037/afb.v3i4.13429
Abo-Kamer AM, Abd-El-salam IS, Mostafa FA, Mustafa A-E-RA, Al-Madboly LA. A promising microbial α-amylase production, and purification from Bacillus cereus and its assessment as antibiofilm agent against Pseudomonas aeruginosa pathogen. Microb Cell Fact. 2023; 22: 141. https://doi.org/10.1186/s12934-023-02139-6
Reji M, Kumar R. Response surface methodology (RSM): An overview to analyze multivariate data. Indian J Microbiol Res. 2022; 9: 241-248. https://doi.org/10.18231/j.ijmr.2022.042
Abdullah R, Ahmad S, Nisar K, Kaleem A, Iqtedar M. Response surface methodology as an approach for optimization of alpha amylase production by using bacterial consortium under submerged fermentation. Kuwait J Sci. 2024; 51: 100220. https://doi.org/10.1016/j.kjs.2024.100220
Adetiloye OA, Solomon BO, Omolaiye JA, Betiku E. Optimization of thermostable amylolytic enzyme production from Bacillus cereus isolated from a recreational warm spring via Box Behnken design and response surface methodology. Microb Cell Fact. 2025; 24: 87. https://doi.org/10.1186/s12934-025-02709-w
Samal KC, Sahoo JP, Behera L, Dash T. Understanding the BLAST (Basic Local Alignment Search Tool) program and a step-by-step guide for its use in life science research. Bhartiya Krishi Anusandhan Patrika. 2021; 36: 55-61. https://doi.org/10.18805/bkap283
Keklik G. Understanding evolutionary relationships and analysis methods through mega software. Int J New Horizons Sci. 2023; 1: 83-90. https://doi.org/10.4135/9781452270142.n1
Sharif S, Shah AH, Fariq A, Jannat S, Rasheed S, Yasmin A. Optimization of amylase production using response surface methodology from newly isolated thermophilic bacteria. Heliyon. 2023; 9. https://doi.org/10.1016/j.heliyon.2023.e12901
Nelson, David L. and MMC. Lehninger principles of biochemistry. 7th ed. New York: W.H. Freeman; 2017.
Kiran S, Singh A, Prabha C, Kumari S, Kumari S. Isolation and characterization of thermostable amylase producing bacteria from hot springs of Bihar, India. Int J Pharma Med Biol Sci. 2018; 7: 28-34. https://doi.org/10.18178/ijpmbs.7.2.28-34
Ozdemir S, Fincan SA, Karakaya A, Enez B. A novel raw starch hydrolyzing thermostable α-amylase produced by newly isolated Bacillus mojavensis SO-10: Purification, characterization and usage in starch industries. Brazilian Arch Biol Technol. 2018; 61: e18160399. https://doi.org/10.1590/1678-4324-2018160399
Rigoldi F, Donini S, Redaelli A, Parisini E, Gautieri A. Engineering of thermostable enzymes for industrial applications. APL Bioeng. 2018; 2. https://doi.org/10.1063/1.4997367
Silva-Salinas A, Rodríguez-Delgado M, Gómez-Treviño J, López-Chuken U, Olvera-Carranza C, Blanco-Gámez EA. Novel Thermotolerant amylase from bacillus licheniformis strain LB04: Purification, characterization and agar-agarose. Microorganisms. 2021; 9: 1857. https://doi.org/10.3390/microorganisms9091857
- چکیده مشاهده شده: 254 بار
- pdf (English) دانلود شده: 152 بار