
Improved production of food-grade hyaluronic acid in recombinant Corynebacterium glutamicum by medium optimization and feeding strategy
Applied Food Biotechnology ,
Vol. 12 Núm. 1 (2025),
4 dey 2025
,
Página 1-14 (e6)
https://doi.org/10.22037/afb.v12i1.46588
Resumen
Abstract
Background and Objective: Hyaluronic acid is extensively used in pharmaceutical, cosmetic, and oral supplementation and nutricosmetic products and has also recently been a candidate for flavor enhancer in the food industry. In this study, Corynebacterium glutamicum ATCC 13032 strain was used for the heterologous production of food-grade hyaluronic acid, and the culture medium and feeding strategy were optimized.
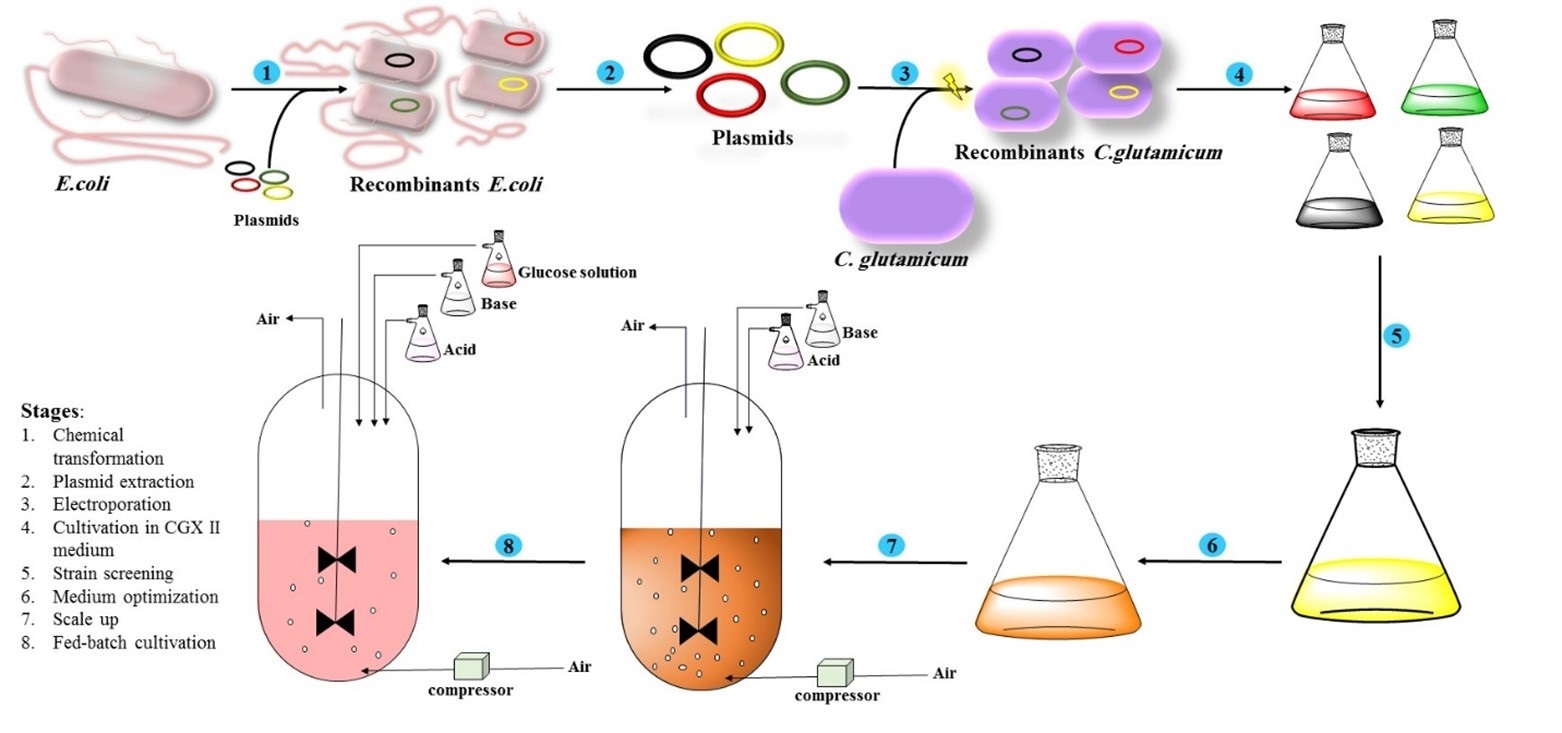
Material and Methods: The propagation of recombinant plasmids was conducted using chemically competent Escherichia coli DH5α, and the extracted plasmids were then transformed into electrocompetent Corynebacterium glutamicum ATCC 13032. A single colony was then transferred into 5 mL of fresh modified CGXII medium supplemented with 50 μg mL⁻¹ kanamycin and incubated for 16-18 h at 30°C. One factor at a time (OFAT) and Taguchi methods were applied to determine the optimal pH and to optimize medium components. Batch, fed-batch, and oxygen-limited fermentation were performed. Hyaluronic acid production was measured using the carbazole and CTAB methods.
Results and Conclusion: The recombinant strain transformed with the two constructs expressing hasA and hasC genes produced the highest amount of hyaluronic acid. The Taguchi L-27 orthogonal array was selected to optimize eleven factors, each at three levels. The results showed that the yield coefficient increased to 71%, and hyaluronic acid production reached 2300 mg L⁻¹. The urea concentration and induction time were considered as significant factors. To enhance hyaluronic acid production, the glucose feeding and oxygen limitation strategies were performed in a bioreactor with a working volume of 4 L. After 48 h, the feeding strategies resulted in a significant increase in the hyaluronic acid yield, reaching roughly 5700 mg L⁻¹. Our results demonstrated that the recombinant Corynebacterium glutamicum containing two main genes of the hyaluronic acid metabolic pathway has a good potential for producing food-grade hyaluronic acid in fed-batch fermentation.
Conflict of interest: The authors declare no conflict of interest.
1. Introduction
In 1934, a type of polysaccharide was discovered, which was later named "halos." This biopolymer consists of N-acetylglucosamine and D-glucuronic acid disaccharide units that are connected alternately by -1, 3 and -1, 4 glycoside linkages to form a high-molecular-weight polysaccha-ride.[1, 2] The molecular weight of this unbranched poly-saccharide is highly variable, ranging from 104 to 4×107 Da.[3]Hyaluronic acid (HA) has some excellent physio-chemical properties such as nontoxicity, nonimmuno-genicity, biocompatibility, and high water absorption capacity, making it an attractive biomolecule for various industrial and biomedical applications.[4]HA has a variety of applications in ophthalmological surgery, cosmetics, regeneration and reconstruction of soft tissues, arthritis rheumatoid and most recently it is introduced as an a drug delivery agent and also use of this polysaccharide have been noticed in food industry such as flavor enhancer which can reduce use of salt up to 10% without affecting saltiness or uses as anti-aging oral supplementation for improving skin physiology, joint health and even muscle strengthening in special cases. Therefore, recently these products such as food-grade HA have been categorized as “nutricosmetics”.[5, 6] Generally, HA with varying mole-cular weights (MW) serves different purposes. High-molecular-weight HA (HMW-HA, ≥1×10⁶ Da) is ideal for joint injections and cartilage repair due to its viscoelasticity and lubrication. Low-molecular-weight HA (LMW-HA, 1×10⁴–1×10⁶ Da) is widely used in cosmetics and products such as juices, jellies, and other food items. Therefore, the demand for hyaluronic acid will grow continuously.[7]
According to a report published in 2024, the global market size for HA was 10.04 billion USD in 2023. With a compound annual growth rate (CAGR) of 7.7%, it is estimated that the market size of HA will reach 16.75 billion USD in 2030.[8]
In the past, HA was produced by extraction from animal sources such as rooster combs and umbilical cords. This method has some disadvantages, including degradation of HA by hyaluronidases, expensive purification methods, and the possibility of viral contamination, which could be considered a serious concern.[9] Therefore, alternative methods such as microbial fermentation are suggested for the production of HA. Microbial fermentation is a process in which the HA is secreted into the culture medium by some bacteria; therefore, the purification costs will significantly decrease. The Streptococcus strains were the first microorganisms used for HA production in bioreactors,[10, 11] However, some concerns, such as endotoxin contaminations, limit their application for medical purposes.[12, 13] Recently, some bacteria such as E. coli, Lactococcus lactis, Bacillus subtilis and Strepto-coccus thermophilus were used for the heterologous production of the HA. These bacteria are categorized as Generally Regarded as Safe (GRAS) microorganisms and are free from any pathogenicity factors and endotoxins.[14] HA is produced by Hyaluronan synthases (HAS) in mammalian and amphibian tissues as well as the cell walls of algae and bacteria.[15, 16] The mammalian genome has three different HAS, and two classes of HAS have been identified in bacteria. HAS, an enzyme typically encoded by the gene named hasA, is a membrane protein that polymerizes HA chains using only Mg²⁺ and two sugar-UDP substrates (UDP-glucuronic acid and UDP-N-acetylglucosamine).[17] The genes involved in HA synthesis are located within the HA operon and consist of hasA, hasB, and hasC genes. Heterologous HA synthesis can be achieved by inserting genes involved in HA production into the genomes of other microorganisms. In some bacteria, hasA is the only essential gene required for HA synthesis.
- glutamicum is a GRAS and hyaluronidase-negative bacterium, making it an ideal candidate for the commercial production of HA [18-20] the genetic engineering tools have facilitated manipulation and insertion of desired genes into C. glutamicum. In this bacterium, other genes, such as glmU, are also involved in the metabolic pathway responsible for HA synthesis. In one branch of this pathway, the expression of hasB and hasC leads to the production of UDP-glucuronic acid. In another branch, glmU encodes a bifunctional enzyme that catalyzes two sequential reactions: first, an acetyltransferase activity that converts glucosamine-1-phosphate into N-acetylglucosamine-1-phosphate, and second, a uridyltransferase activity that converts N-acetylglucosamine-1-phosphate into UDP-N-acetylglucosamine. Finally, hasA, which encodes hyaluronan synthase, serves as the intersection point of these two branches and polymerizes HA from UDP-N-acetylglucosamine and UDP-glucuronic acid. HA synthesis by C. glutamicum was first reported in 2014 with a yield of 1241 mg L⁻¹ after 120h of fermentation.[19] In a subsequent study, various genetic engineering approaches and strategies were employed, including the use of strong promoters, different plasmid constructs, and the evaluation of various induction times, to enhance high-titer biosynthesis of HA in C. glutamicum. The findings revealed that the strain harboring the artificial ssehasA gene derived from Streptococcus equisimilis with C. glutamicum codon preference and the hasB gene, utilizing the Ptac inducible promoter, produced HA within the range of 1.77 to 2.23 g L⁻¹. Under optimal conditions, the production reached 5.25 g L⁻¹ while under non-pH control HA titer increased significantly and reached an impressive 8.3 g L⁻¹ at 48h fermentation.[21] The study on this strain continued as engineered C. glutamicum achieved high-titer HA production. A genome-scale metabolic model was utilized to identify genetic interventions through flux balance analysis. The focus was enhancing the HA biosynthesis pathway while attenuating the glycolysis pathway and knocking out competing pathways. Various genetic strategies resulted in a surprisingly high HA titer of 28.7 g L⁻¹ in the engineered C. glutamicum.[22] This outcome demonstrates the power of molecular approaches compared to traditional fermentation strategies. Another novel strategy for enhancing HA production in C. glutamicum focuses on cell morphology through a well-designed dual-valve regulation system. This system comprises two modules: a transporter module featuring a strong constitutive promoter (Ptuf) and an arabinose transport protein, and a morphology-tuning module with an arabinose-inducible weak promoter (PBAD) and a cell-division-related gene. This approach enables fine-tuning of cell morphology, increasing cell length by 1.87-fold and cell membrane size by 2.08-fold, ultimately achieving an HA titer of 16.0 g L⁻¹. This represents a 1.6-fold improvement in yield compared to previous studies on morphology-engineered strains, underscoring the potential of this strategy for enhancing HA production [23].
In this study, four recombinant expression plasmids were introduced into C. glutamicum, and hyaluronic acid (HA) production was analyzed for individual plasmids and their combinations to identify the most effective expression vectors for maximizing HA production. The goal of this research is to enhance HA titer by developing an optimized culture medium and refining the medium and fermentation conditions. To achieve this, the Taguchi design of experiments was employed to optimize a chemically defined medium for HA synthesis in flasks. Ultimately, HA production in the optimized medium was evaluated under controlled conditions, including oxygen limitation and glucose feeding, in a 5-liter fermenter.
- Materials and Methods
2.1 Microorganisms and plasmids
Escherichia coli DH5α was used for propagation of recombinant plasmids. Corynebacterium glutamicum ATCC 13032 and recombinant plasmids were kindly donated by Josef Altenbuchner from The University of Stuttgart, Germany, and were used for HA production. The recombinant plasmids containing the genes involved in the HA production named pAC (harboring hasA and hasC), pACB (harboring hasA, hasC and hasB in order in operon), pA (harboring just hasA) and pAGC (harboring hasA, glmU and hasC in order).[19]
2.2 Media and Cultivation
- coli was cultivated in Luria-Bertani (LB) medium containing 10 g L⁻¹ tryptone, 10 g L⁻¹ NaCl, and 5 g L⁻¹ yeast extract supplemented with 50 μg ml-1 kanamycin. For LB-agar preparation, 15 g L⁻¹ agar was added. Recombinant C. glutamicum was cultivated in a modified CGXII medium for HA production as follows. Solutions were prepared separately and then mixed. The first solution was prepared by dissolving 5 g urea, 5 g (NH4)2SO4, 1 g K2HPO4, and 1 g KH2PO4 in 800 ml distilled water. To prepare the second solution, 250 mg MgSO4 and 10 mg CaCl2 dissolved in 50 ml distilled water. The third solution was prepared by dissolving 10 g of glucose in 50 ml distilled water. All the solutions were autoclaved after preparation, except for the glucose solution, which was sterilized by filtration. After cooling, the solutions were mixed. The trace element solution was prepared and sterilized, and 1 ml was added to the medium as mentioned above. A vitamin solution containing 1mg/ml biotin was prepared, the filter was sterilized, and 0.2 ml was added to the primary medium. The final volume was adjusted to 1 liter with sterile distilled water [19, 24].
2.3 Competent cells preparation and transformation
- coli competent cells were prepared using a chemical method using CaCl2 and transformed by a heat shock procedure at 42°C.[25] For electrocompetent cell preparation, a single colony of C. glutamicum was transferred into 5 ml brain-heart infusion (BHI) broth medium and incubated for 18 hours at 30°C and 180 rpm. Then, 2 ml of bacterial suspension was inoculated into a 100 ml electroporation medium containing 37 g l-1 BHI, 0.1% v/v tween 80, 25 g l-1 glycine, and cultivated at 30 ᵒC and 180 rpm to reach OD600=0.8. The bacteria were then centrifuged at 3000 ×g for 15 min at 4ᵒC, and the precipitate was washed with 20% v/v glycerol. The centrifugation and glycerol washing steps were repeated three times. Finally, the pellet was resuspended in 1 ml of 15% (v/v) glycerol and stored at -70°C. [26]
Electroporation was carried out using Gene Pulser II (Bio-Rad) as follows: first, 100 ng of supercoiled plasmid DNA was mixed gently with 100 μl electrocompetent cells and transferred into a 0.2 cm (2 mm) cuvette. The electroporator was set to 2.5 kV, 25 μF, and 200 Ω. Immediately after the pulse,1 ml BHI medium was added to the bacteria and incubated for 6 min at 46°C followed by 1 h incubation at 30°C. Finally, the bacteria were plated on solidified medium supplemented with 50 mL-1 kanamycin.[19, 26]
2.4 Fermentation condition in shake flask
For pre-culture preparation, a single colony was transferred into 5 mL fresh modified CGXII medium supplemented with 50 μg mL-1 kanamycin and incubated for 16-18 h at 30°C and 180 rpm. One mL of overnight culture was inoculated into 25 mL fresh medium supplemented with 50 μg ml-1 kanamycin. The induction of the bacteria was carried out using 1 mM IPTG (Isopropyl β-D-1-thiogalactopyranoside) at different OD600 initiating from OD600= 0.5. IPTG is used as a molecular mimic of allolactose to induce the expression of our genes, which are under the control of the lac promoter. All flasks were incubated for 24-120 h at 30°C and 180 rpm. The pH, glucose consumption, HA, OD600, and biomass production were measured at different time intervals. In some cases, if necessary, 4% glucose was also added after 48 h.
2.5 Hyaluronic acid quantification
HA production was measured using carbazole and CTAB methods.[27, 28] For this purpose, the bacterial suspension was centrifuged and the supernatant was used for HA assay. In the carbazole method, 1 ml of supernatant was mixed with 2 ml of absolute ethanol and incubated at -20°C overnight. The samples were then centrifuged for 30 min at 3500 g for HA precipitation. After that, the pellet was dissolved in 1 ml deionized water, and carbazole assay was performed as follows: 50 μl sample was added to a 96 well plate and 200 μl solution A (25 mM L⁻¹ sodium tetraborate in sulfuric acid) was added to it. The mixture was incubated for 15 minutes in boiling water and 10 minutes on ice. Then 50 μl solution B (0.125% carbazole in absolute ethanol (v/w)) was added to each well and incubated in boiling water for 10 min. Finally, the absorbance was read at OD540 nm by an ELISA reader.[27] The calibration curve was prepared using different HA concentrations (25, 50, 250, 500, 750, and 1000 mg L⁻¹ ) and a linear equation was used for the calculation of the HA amount.
The CTAB method added 50 μl HA samples to a 96-well plate. Then 50 μl acetate buffer (0.2 M sodium acetate, 0.15 M sodium chloride, pH 6) was added to each well and incubated at 37°C for 10 min. After that, 100 μl CTAB solution (25 g L⁻¹ CTAB dissolved in 2% NaOH) was added to the well, and the absorption was read at 600 nm after 10 min by an ELISA reader.[28]
2.6 Glucose assay
Glucose concentration was determined using an enzymatic kit (Pars Azmoon Co). The calibration curve was prepared for seven different concentrations (0.25, 0.5, 1, 2, 2.5, 3.5, and 4.5 g L⁻¹ ) of glucose, and a linear equation was used to calculate the amounts.
2.7 Cell growth measurement
Cell growth was monitored by measuring optical density at 600 nm and cell dry weight.
2.8 Statistical methods
One factor at a time (OFAT) method was applied to find the best pH (6, 7, and 8). Taguchi method was carried out for optimization of medium components (phosphate buffer (K2HPO4, KH2PO4), Ca (NO3)2, (NH4)2SO4, MgSO4, soy protein acid hydrolysate, biotin, trace elements, glucose, citric acid, urea) and induction time (Table 1). L-27 orthogonal array with eleven factors in three levels was selected to design experiment (Table 2). Experiment design and analysis were performed by Qualitek-4 (version 4.82.0) software.
2.9 Batch, fed-batch, and oxygen-limited fermentation in a 5L fermenter
A loop from the fresh plate was picked up, transferred into the 20 ml modified CGX II medium, and incubated at 30 °C overnight. Then, 400 ml modified CGX II medium was inoculated with 20 ml overnight culture and incubated for 10 h to reach OD600nm= 4 to 5. The overnight culture was then applied for vessel inoculation. The optimized medium was prepared in 4-L volume and batch, fed-batch, and oxygen-limited was performed at 30 °C, pH controlled at 7, and initial OD600nm adjusted at 0.4-0.5. Batch fermentation was performed for 24 h with 200-600 rpm agitation rate and 20-40 percent dissolved oxygen. Oxygen-limited fermentation was performed for 24 h with a 200 rpm agitation rate, while dissolved oxygen was controlled between 0-5 percent.[21]
Fed-batch fermentation was performed for 48 h with 200-600 rpm agitation rate and dissolved oxygen was controlled between 20-40 percent. After 18 h, 400 ml feeding solution containing glucose 60%, (NH4)2SO4 1.5 g L⁻¹, MgSO4 5g L⁻¹, yeast extract 20 g L⁻¹, IPTG 1 mM, kanamycin 50 mg L⁻¹ and trace elements 1ml L⁻¹ was prepared and added to the vessel. The feeding rate was adjusted to 15 ml h⁻¹.
- Results and Discussion
3.1 Effect of different genes involving metabolic pathway of HA
The yield of HA production by four constructed vectors was pAC > pACB> pA > pAGC. The yield of HA produced by the pA construct harboring hasA gene, showed that the hasA was the most important gene in this process (Fig. 1 and 2). The comparison of the HA production by C. glutamicum transformed with the pA construct and the wild-type strain demonstrated that the HA synthase expression was required for high yield of HA production. In fact, the limiting step in HA production by C. glutamicum was HA synthase. Our results concur with previous studies on HA synthesis by gram-negative and positive bacteria.[12, 13, 29, 30]
The Comparison of the HA production by pA and pAC recombinant constructs (containing hasA and hasC genes, respectively) revealed that the hasC gene had only a minor impact on enhancing the HA yield. The presence of the hasC gene resulted in a 12% increase in HA production (Fig. 1 and 2). This result confirmed the previous findings that C. glutamicum contains a pool of precursors for HA synthesis.[31]
Overexpression of glmU gene decreased the HA yield and the cell concentration by 3.5 and 18-fold, respectively (Figures 1 and 2). The glmU gene encodes for a uridyltransferase enzyme that produces UDP-GlcNAc. It seems that increased precursor concentrations inhibit bacterial growth; therefore, a high level of UDP-GlcNAc results in a reduction in the cell concentration and HA synthesis.[29] To compare our results from C. glutamicum with another alternative host for HA production, we can look at findings reported by Zichao Mao and his colleagues, who worked with Escherichia coli, a gram-negative bacterium. They transformed several genes, including uridine diphosphate-glucose dehydrogenase from E. coli K5 and pmHAS from Pasteurella multocida, which are key genes for HA production in E. coli. Their results showed a yield of 0.5 g L⁻¹ in shaking flasks and approximately 2.0–3.7 g L⁻¹ in 1 L fed-batch fermenters[30].
Based on these results, we can conclude that C. glutamicum is a better option than E. coli, as it requires only one gene to produce HA and does not contain endotoxins in the final product. Additionally, the minimum yield of HA production in C. glutamicum is higher than that of E. coli. In another study, Naoki Izawa and his colleagues attempted to produce HA in Streptococcus thermophilus. They reported a maximum titer of 1.2 g L⁻¹ with the co-expression of hasA and hasB, which is similar to the minimum yield in C. glutamicum. Furthermore, Lactococcus lactis was chosen for HA production due to its status as a food-grade bacterium. The researchers claimed that HA produced by L. lactis has significant potential for applications in the food and biomedical industries[29]. However, the maximum titer reported in their study was only 0.65 g L⁻¹, which is again lower than the minimum HA production obtained from C. glutamicum. Another alternative host for HA production is Bacillus subtilis. Bill Widner and his colleagues transformed several genes, including hasA, tauD, and gcaD, reporting yields of over 1 g L⁻¹ [13]. Although C. glutamicum has a higher titer compared to the results from this study, B. subtilis has shown great potential, and many researchers are conducting studies on it. Today, some manufacturers are using B. subtilis as an alternative host for industrial HA production. We believe that C. glutamicum, along with Bacillus subtilis, represents the best options for producing hyaluronic acid. Our results, along with other studies, indicate that C. glutamicum is one of the bacteria with a high titer of HA production, slightly lower than the native HA producer, Streptococcus zooepidemicus.
In industrial production, an important challenge arises when working with plasmids and recombinant strains: plasmid instability. This issue occurs when recombinant strains lose the plasmid or experience a decrease in copy number over generations, leading to unstable expression and a reduction in the titer of products like HA. C. glutamicum is no exception to this problem. One approach to address this challenge is developing and using integrative plasmids, which integrate the gene of interest into the genome of C. glutamicum. This method resolves most stability issues. Additionally, some studies on C. glutamicum have identified a gene named cgR_0322, which is involved in the response to plasmid introduction and plasmid structural instability. Disrupting this gene may enhance plasmid retention and expression of harbored genes, thereby broadening the bacterium’s suitability as an industrial production host.[32]
3.2 Effects of initial pH on HA production
A one-factor-at-a-time method was applied to find the best pH for HA production in the culture medium. As expected, neutral pH was the best pH for HA production.[18] The high pH causes sedimentation and turbidity in the medium due to the reduced solubility of some components, such as phosphate salts and proteins. On the other hand, low pH inhibits bacterial growth and HA production by causing cellular stress and disrupting bacterial membrane integrity. Additionally, the growth of C. glutamicum is typically accompanied by the secretion of acidic byproducts into the medium. Therefore, starting with a pH around neutral is better to avoid extreme decreases in pH during fermentation. For these reasons, a neutral pH was chosen.
3.4. Optimization of the HA production by Taguchi method, data analysis by Qualitek-4 software
The ANOVA table was generated using Qualitek-4 software based on the data obtained. According to the analysis, urea concentration (F-ratio = 31.658) emerged as the most significant factor influencing hyaluronic acid (HA) production. Induction time also demonstrated substantial importance, further validating its role as a critical parameter. The ANOVA table summarizes the detailed effects of each factor on HA production (Table 3).
The results highlight urea (F = 31.66, 43.1% contribution) and induction time (F = 16.60, 22.0% contribution) as the dominant factors, collectively accounting for over 65% of the total variance. Their high F-values, which are well above the significance threshold, underscore their statistical and practical relevance. Soy protein acid hydrolysate (F = 6.67, 8.0%) and vitamin complex (F = 5.66, 6.6%) exhibited moderate influence, likely by supporting microbial growth and precursor synthesis. MgSO₄ (F = 3.45, 3.4%) and citric acid (F = 2.39, 2.0%) showed minor but measurable effects. Remaining factors, such as phosphate buffer, glucose, and trace elements, contributed negligibly (F < 1, % < 1%), indicating minimal impact under the tested conditions. These findings prioritize urea concentration and induction timing as key variables for optimizing HA yield, while deemphasizing non-significant factors. The model explained approximately 85% of the total variability (14.7% unexplained error). The impacts of various factors on the response values were analyzed using signal-to-noise ratio and the plots were drawn for each factor (Fig. 3). The graphs display the maximum and minimum responses for each level, suggesting the best level for each factor as well as optimum condition.
3.4 The effects of phosphate buffer, ammonium sulfate and trace elements
The concentration of phosphate buffer components had no significant impact on HA production (Fig. 3-a). Under all conditions, the pH of the medium dropped within the first 8 hours. Furthermore, increasing the buffer concentration also had no major effects (Table 4). Similarly, ammonium sulfate, as a mineral source of nitrogen, had no significant impact on HA production; however, the second level of ammonium sulfate was more effective for HA production (Fig. 3-b). Our results also demonstrated that trace elements had no major effects on HA production (Fig. 3-f).
3.5 The effect of calcium nitrate
As shown in Fig. 3-c, increasing the amount of calcium nitrate as a source of calcium ion marginally decreased the HA production.
3.6 The effects of soy protein lysate
In this study, soy protein lysate was used as a source of amino acids and organic nitrogen. The maximum amount of HA was produced in the second level, while the HA production in the first and third levels was lower than the second level (Fig 3-d). As can be deduced from Table 4, using soy protein acid hydrolysate in the highest amounts had deleterious impacts on the HA yield.
3.7 The effect of biotin
Biotin was added to the culture medium in a form of the B complex vitamin batch. As shown in Fig. 3-e, increasing the biotin concentration from the first level to the second level increased the HA production, whereas increasing the biotin concentration from the second level to the third level decreased the production.
3.8 The effects of initial glucose concentration
UDP-GlcNAc and D-glucuronic acid, derived from glucose and produced in the carbon pathways, are the major components for the HA backbone synthesis. In this study, the glucose concentrations were considered high, while the bacterial concentration had no limitation. When the initial glucose concentration in the first level was considered high, the further increase in the glucose concentration slightly affected the HA yield. When the glucose concentration in the first level was considered low, the increase in the glucose concentration had stronger impacts on the HA synthesis (Fig. 3-g). In low glucose condition, a slight increase in the glucose concentration increased the HA production, whereas in high initial glucose concentrations, the bacteria had unrestricted access to the carbon source, so a further increase in glucose concentration had no effects. Also, according to the results of Table 4, it can be concluded that the impact of initial glucose concentration on HA synthesis could be significant at low initial glucose concentrations.
3.9 The effect of citric acid
The effects of citrate on the growth of C. glutamicum were also investigated. The presence of citrate in the culture medium increased the expression levels of certain enzymes involved in TCA cycle and regulation of the central metabolism in C. glutamicum.[33] Fig. 3-h results can also confirm that increasing the citrate concentration boosted precursor synthesis and HA production.
3.10 The effect of MgSO4
Magnesium sulfate is the fifth important cofactor for Hyaluronan synthase. The maximum amount of HA was achieved at 200 mg L⁻¹ concentration of MgSO4. Subsequent increase in MgSO4 concentration decreased the HA production. It seems that MgSO4 at high quantities blocks enzymes involved in the carbon cycle and HA precursor synthesis, such as phosphoenolpyruvate carboxykinase and pyruvate carboxylase that use Mn2+ as cofactor.[34] Actually, Mg2+ has a similar atomic radius to Mn2+, so at high concentrations, it can bind to the enzymes instead of Mn2+ and reduce their activities.[35] The results of Fig. 3-i can be interpreted as the presence of high quantities of MgSO4 in the culture media might disrupt enzymes involved in the carbon cycle and HA precursor synthesis that use Mn2+ as a cofactor, such as phosphoenolpyruvate carboxykinase and pyruvate carboxylase.
3.11 The effects of induction time
This experiment demonstrated that adding an inducer at different time points had major effects on HA production. The addition of IPTG as an inducer at the low optical density of bacteria induced HA production and decreased bacterial growth (Fig. 2). The reduction in bacterial growth could be due to the competition between HA production and cell wall synthesis.[16, 36] On the other hand, the induction of bacteria at higher OD had no effects on HA production (Fig. 3-j). Actually, when bacteria are growing, the precursors for cell wall synthesis are present in the cells, and the addition of IPTG at this time results in the production of HA. On the contrary, when the cells are in the late stages of growth or the last log phase, there are no precursors for HA production; therefore, adding IPTG does not affect the HA synthesis. Furthermore, in the late stages of bacterial growth, the metabolic pathways switch to biomass production and adding IPTG cannot change the pathways for HA production.
3.12 The effects of urea
The urea concentration was another variable that was subjected to the optimization process. The results in Table 2 revealed that glucose was consumed entirely in some conditions in which the pH of the culture media was between 6-7. In other runs, the pH of the culture media was acidic between 4-5. It seems glucose depletion leads to urea hydrolysis and NH3+ production, raising pH to neutral level and stimulating HA synthesis (Fig. 3-k). In some runs shown in Table 2, glucose was not fully consumed (runs 9, 10, 18, and 19), urea hydrolysis was suppressed, and pH remained in the acidic range. A possible known mechanism for the effect of urea on HA production is supported by research identifying the urea uptake system (urtABCDE operon) and urease genes (ureABCEFGD), which are regulated by the global nitrogen regulator AmtR under nitrogen-limiting conditions. Studies have shown that under nitrogen limitation, the synthesis of urease subunits increases, making urea utilization critical for nitrogen supply.[37]This confirms the impact of urea on metabolic flux and potentially glucose uptake, aligning with our observations. Urea hydrolysis produces ammonia, which is assimilated into nitrogen metabolism via glutamine synthetase (GlnA).[38] GlnA converts ammonia into glutamine, which then donates an amino group for the formation of glucosamine-6-phosphate. This compound serves as a precursor in the metabolic pathway leading to UDP-N-acetylglucosamine (UDP-GlcNAc) synthesis. UDP-GlcNAc is one of the two sugar-UDP substrates required for HA polymerization
3.13 Verification test
The optimum levels of the factors should be experimentally confirmed. The qualiteq-4 software predicted the maximum HA concentration in the range of 1299 to 3798 mg L⁻¹ with a 95% confidence interval. The verification test indicated that the HA concentration reached 2300 mg L⁻¹, which was in line with the predicted range. HA production remarkably increased by the Taguchi experimental design, and it can be concluded that the statistical approach was efficient.
3.14 Fermentation conditions
The optimal condition was tested in a 5-L fermenter and showed similar results. The HA production reached 1.8 g L⁻¹ after 18 h (Fig. 4). At pH 7, glucose was consumed entirely, and the production decreased. On the other hand, in the acidic pH of the culture media, the bacteria could not consume glucose, and HA production decreased. The results indicated glucose depletion was a critical limiting step in HA synthesis. Actually, at neutral pH, the HA was produced when glucose was not totally consumed.
In our study, glucose feeding had a major impact on HA synthesis. The feeding was started when the initial glucose was completely consumed. After 48 h, the HA production reached 5.3 g L⁻¹ (Fig. 5). Glucose feeding maintained bacterial growth; therefore, there were no limitations for HA production. When bacteria are in the growth phase, the precursors necessary for HA production are available and promote both HA production and cell division.[36]
In the fermentation process for the production of HA, an important challenge to consider is oxygen transfer limitation. HA, a large polysaccharide and water absorber, increases the viscosity of the medium. This increased viscosity reduces oxygen availability and dissolved oxygen levels, which negatively affects cell growth and HA production. While higher aeration may partially address the issue, research on Streptococcus zooepidemicus indicates that the controlled use of the hyaluronidase enzyme, which converts high molecular weight HA into lower molecular
weight forms, can enhance oxygen transfer and improve dissolved oxygen levels.[39] This approach helps mitigate the oxygen limitation problem and enhances the oxygen transfer rate, leading to an increase in the titer of HA production, although the final HA product will have a lower molecular weight. This approach could likely be effective for C. glutamicum as well, and using it could resolve this issue for industrial production.
- Conclusion
The results demonstrated that the hasA gene is crucial for recombinant HA production in C. glutamicum. While some bacteria produce HA precursors like N-acetylglucosamine and D-glucuronic acid, they cannot assemble HA without the hasA gene. Introducing hasA enables HA production, though genes like glmU may negatively impact HA yield by disrupting metabolic pathways. Medium optimization using the Taguchi method and ANOVA in this study, identified urea concentration and induction time as significant factors for HA production with an F-ratio more than the F critical value (6.9443). Urea, as an organic nitrogen source, adjusts pH and enhances glucose uptake, leading to higher HA production when glucose is fully consumed. Higher urea concentrations prevent pH decline by producing ammonium, maintaining pH at 6.5–7.5, which is optimal for glucose consumption and HA production. Conversely, low pH reduces glucose uptake and HA yield.
Induction time also significantly influenced HA production. Early induction By IPTG (at low bacterial concentration) directed precursors toward HA synthesis, while late induction (at high OD) inhibited HA production due to competition between bacterial growth and HA synthesis. Since HA production is growth-associated, maintaining bacteria in the log phase increased HA yield. Glucose feeding and pH adjustment in the fermenter further enhanced HA production by preventing entry into the stationary phase and maintaining pH around 7. In conclusion, C. glutamicum with the hasA gene can produce high HA yields when grown in urea-enriched medium, induced early, and fed glucose to sustain growth and pH. Therefore, it could be a candidate as an alternative host for industrial HA production.
- Acknowledgements
This research was financially supported by the National Institute of Genetic Engineering and Biotechnology (NIGEB) in Tehran. Iran (project #643). We greatly appreciate it.
- Conflict of Interest
The authors have no conflicts of interest to declare. All co-authors have seen and agree with the contents of the manuscript, and there is no financial interest to report. We certify that the submission is original work and is not under review at any other publication.
- Author Contributions
All authors participated in project administration and writing of the first draft of the manuscript, providing critical revision and editing. All authors approved the final version of the manuscript.
- Corynebacterium glutamicum
- Fed-batch fermentation
- Hyaluronic acid
- Optimization
- Taguchi experimental design
Cómo citar
Citas
Saravanakumar K, Park S, Santosh SS, Ganeshalingam A, Thiripuranathar G, Sathiyaseelan A, et al. Application of hyaluronic acid in tissue engineering, regenerative medicine, and nanomedicine: A review. International Journal of Biological Macromolecules. 2022;222:2744-60. https://doi.org/10.1016/j.ijbiomac.2022.10.055
Meyer K, Palmer JW. The polysaccharide of the vitreous humor. Journal of Biological Chemistry. 1934;107(3):629-34. https://doi.org/10.1016/S0021-9258(18)75338-6
Lee BM, Park SJ, Noh I, Kim C-H. The effects of the molecular weights of hyaluronic acid on the immune responses. Biomaterials Research. 2021;25(1):27. https://doi.org/10.1186/s40824-021-00228-4
Bayer IS. Hyaluronic acid and controlled release: A review. Molecules. 2020;25(11):2649. https://doi.org/10.3390/molecules25112649
Graciela CQ, Jose Juan EC, Gieraldin CL, Xochitl Alejandra PM, Gabriel AA. Hyaluronic Acid-Extraction Methods, Sources and Applications. Polymers (Basel). 2023;15(16). https://doi.org/10.3390/polym15163473
Hu J, Chen Z, Huang X, Yan Z, Li Y, Zhu Y, et al. Hyaluronic acid applied as a natural flavor enhancer and its mechanism exploration. Food Bioscience. 2023;55:102969. https://doi.org/10.1016/j.fbio.2023.102969
Qiu Y, Ma Y, Huang Y, Li S, Xu H, Su E. Current advances in the biosynthesis of hyaluronic acid with variable molecular weights. Carbohydrate polymers. 2021;269:118320. https://doi.org/10.1016/j.carbpol.2021.118320
Hyaluronic Acid Market Size, Share & Trends Analysis Report By Application (Dermal Fillers, Osteoarthritis,Ophthalmic, Vesicoureteral Reflux), And Segment Forecasts, 2024 - 2030. https://www.grandviewresearch.com/industry-analysis/hyaluronic-acid-market#; 2024.
Murado M, Montemayor M, Cabo M, Vázquez J, González M. Optimization of extraction and purification process of hyaluronic acid from fish eyeball. Food and bioproducts processing. 2012;90(3):491-8. https://doi.org/10.1016/j.fbp.2011.11.002
Chong BF, Nielsen LK. Aerobic cultivation of Streptococcus zooepidemicus and the role of NADH oxidase. Biochemical engineering journal. 2003;16(2):153-62. https://doi.org/10.1016/j.fbp.2011.11.002
Krahulec J, Krahulcová J. Increase in hyaluronic acid production by Streptococcus equi subsp. zooepidemicus strain deficient in β-glucuronidase in laboratory conditions. Applied microbiology and biotechnology. 2006;71:415-22. https://doi.org/10.1007/s00253-005-0173-9
Chien L-J, Lee C-K. Hyaluronic acid production by recombinant Lactococcus lactis. Applied microbiology and biotechnology. 2007;77:339-46. https://doi.org/10.1007/s00253-007-1153-z
Widner B, Behr R, Von Dollen S, Tang M, Heu T, Sloma A, et al. Hyaluronic acid production in Bacillus subtilis. Applied and environmental microbiology. 2005;71(7):3747-52. https://doi.org/10.1128/AEM.71.7.3747-3752.2005
Shikina E, Kovalevsky R, Shirkovskaya A, Toukach PV. Prospective bacterial and fungal sources of hyaluronic acid: A review. Computational and Structural Biotechnology Journal. 2022;20:6214-36. https://doi.org/10.1016/j.csbj.2022.11.013
DeAngelis P. Hyaluronan synthases: fascinating glycosyltransferases from vertebrates, bacterial pathogens, and algal viruses. Cellular and Molecular Life Sciences CMLS. 1999;56:670-82. https://doi.org/10.1007/s000180050461
O'Regan M, Martini I, Crescenzi F, De Luca C, Lansing M. Molecular mechanisms and genetics of hyaluronan biosynthesis. International Journal of Biological Macromolecules. 1994;16(6):283-6. https://doi.org/10.1016/0141-8130(94)90056-6
Weigel PH. Hyaluronan synthase: the mechanism of initiation at the reducing end and a pendulum model for polysaccharide translocation to the cell exterior. International Journal of Cell Biology. 2015;2015(1):367579. https://doi.org/10.1155/2015/367579
Eggeling L, Bott M. Handbook of Corynebacterium glutamicum: CRC press; 2005. https://doi.org/10.1201/9781420039696
Hoffmann J, Altenbuchner J. Hyaluronic acid production with Corynebacterium glutamicum: effect of media composition on yield and molecular weight. Journal of applied microbiology. 2014;117(3):663-78. https://doi.org/10.1111/jam.12553
Crater DL, Van de Rijn I. Hyaluronic Acid Synthesis Operon (has) Expression in Group A Streptococci (∗). Journal of Biological Chemistry. 1995;270(31):18452-8. https://doi.org/10.1074/jbc.270.31.18452
Cheng F, Gong Q, Yu H, Stephanopoulos G. High‐titer biosynthesis of hyaluronic acid by recombinant Corynebacterium glutamicum. Biotechnology journal. 2016;11(4):574-84. https://doi.org/10.1002/biot.201500404
Cheng F, Yu H, Stephanopoulos G. Engineering Corynebacterium glutamicum for high-titer biosynthesis of hyaluronic acid. Metabolic engineering. 2019;55:276-89. https://doi.org/10.1016/j.ymben.2019.07.003
Yuan S, Zheng Y, Du Y, Song M, Sun CC, Cheng F, Yu H. Fine-tuning the cell morphology of Corynebacterium glutamicum via dual-valve regulation for enhanced hyaluronic acid production. Biotechnology Notes. 2023;4:135-45. https://doi.org/10.1016/j.biotno.2023.12.003
Keilhauer C, Eggeling L, Sahm H. Isoleucine synthesis in Corynebacterium glutamicum: molecular analysis of the ilvB-ilvN-ilvC operon. Journal of bacteriology. 1993;175(17):5595-603. https://doi.org/10.1128/jb.175.17.5595-5603.1993
Sanbrook J, Fritsch E, Maniatis T. Molecular cloning: a laboratory manual. Cold Spring Harbor, NY, Cold Spring Harbor Laboratory. 1989;11:31.
Van der Rest M, Lange C, Molenaar D. A heat shock following electroporation induces highly efficient transformation of Corynebacterium glutamicum with xenogeneic plasmid DNA. Applied Microbiology and Biotechnology. 1999;52:541-5. https://doi.org/10.1007/s002530051557
Bitter T, Muir HM. A modified uronic acid carbazole reaction. Analytical biochemistry. 1962;4(4):330-4. https://doi.org/10.1016/0003-2697(62)90095-7
Chen Y-H, Wang Q. Establishment of CTAB Turbidimetric method to determine hyaluronic acid content in fermentation broth. Carbohydrate polymers. 2009;78(1):178-81. https://doi.org/10.1016/j.carbpol.2009.04.037
Izawa N, Serata M, Sone T, Omasa T, Ohtake H. Hyaluronic acid production by recombinant Streptococcus thermophilus. Journal of bioscience and bioengineering. 2011;111(6):665-70. https://doi.org/10.1016/j.jbiosc.2011.02.005
Mao Z, Shin H-D, Chen R. A recombinant E. coli bioprocess for hyaluronan synthesis. Applied microbiology and biotechnology. 2009;84:63-9. https://doi.org/10.1007/s00253-009-1963-2
De Bruyn F, Maertens J, Beauprez J, Soetaert W, De Mey M. Biotechnological advances in UDP-sugar based glycosylation of small molecules. Biotechnology Advances. 2015;33(2):288-302. https://doi.org/10.1016/j.biotechadv.2015.02.005
Kitade Y, Okino S, Gunji W, Hiraga K, Suda M, Suzuki N, et al. Identification of a gene involved in plasmid structural instability in Corynebacterium glutamicum. Applied microbiology and biotechnology. 2013;97:8219-26. https://doi.org/10.1007/s00253-013-4934-6
Polen T, Schluesener D, Poetsch A, Bott M, Wendisch VF. Characterization of citrate utilization in Corynebacterium glutamicum by transcriptome and proteome analysis. FEMS microbiology letters. 2007;273(1):109-19. https://doi.org/10.1111/j.1574-6968.2007.00793.x
Baly DL, Keen CL, Hurley LS. Pyruvate carboxylase and phosphoenolpyruvate carboxykinase activity in developing rats: effect of manganese deficiency. The Journal of nutrition. 1985;115(7):872-9. https://doi.org/10.1093/jn/115.7.872
Kayestha R, Hajela K. ESR studies on the effect of ionic radii on displacement of Mn2+ bound to a soluble β-galactoside binding hepatic lectin. FEBS letters. 1995;368(2):285-8. https://doi.org/10.1016/0014-5793(95)00673-W
de Oliveira JD, Carvalho LS, Gomes AMV, Queiroz LR, Magalhães BS, Parachin NS. Genetic basis for hyper production of hyaluronic acid in natural and engineered microorganisms. Microbial cell factories. 2016;15:1-19. https://doi.org/10.1186/s12934-016-0517-4
Beckers G, Bendt AK, Krämer R, Burkovski A. Molecular identification of the urea uptake system and transcriptional analysis of urea transporter-and urease-encoding genes in Corynebacterium glutamicum. Journal of Bacteriology. 2004;186(22):7645-52. https://doi.org/10.1128/jb.186.22.7645-7652.2004
Burkovski A. Ammonium assimilation and nitrogen control in Corynebacterium glutamicum and its relatives: an example for new regulatory mechanisms in actinomycetes. FEMS Microbiology Reviews. 2003;27(5):617-28. https://doi.org/10.1016/S0168-6445(03)00067-6
Liu L, Du G, Chen J, Wang M, Sun J. Influence of hyaluronidase addition on the production of hyaluronic acid by batch culture of Streptococcuszooepidemicus. Food chemistry. 2008;110(4):923-6. https://doi.org/10.1016/j.foodchem.2008.02.082
- Resumen ##plugins.themes.ojsPlusA.frontend.article.viewed##: 856 ##plugins.themes.ojsPlusA.frontend.article.times##
- pdf (English) ##plugins.themes.ojsPlusA.frontend.article.downloaded##: 319 ##plugins.themes.ojsPlusA.frontend.article.times##