
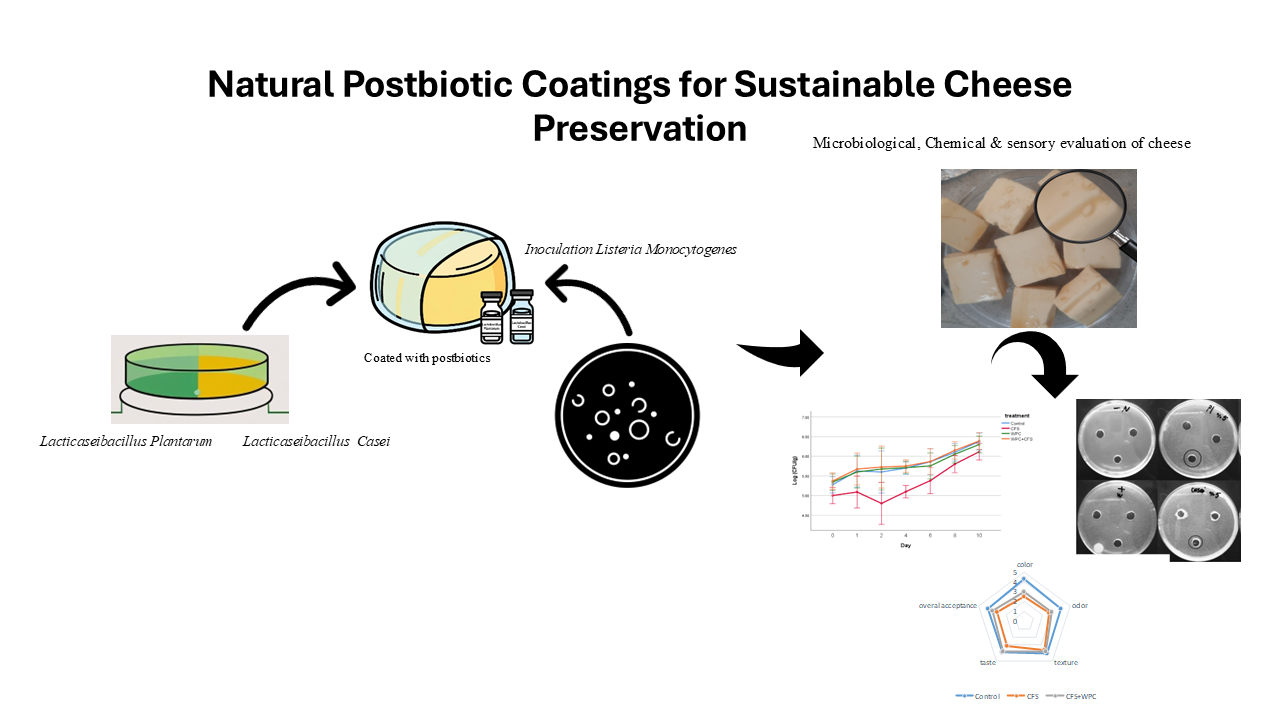
Cutting-Edge Cheese Crafting: Exploring the Benefits of Postbiotics Coating in Pasteurized Cheese Production
بیوتکنولوژی غذایی کاربردی,
دوره 12 شماره 1 (2025),
4 ژانویهٔ 2025
,
صفحه 1-12 (e25)
https://doi.org/10.22037/afb.v12i1.50103
چکیده
Background and Objective: Cheese is one of the major dairy products with high nutritional value, but its susceptibility to microbial growth and early spoilage remains a challenge for the dairy industry. While chemical additives are widely applied to control microbial contamination, increasing awareness of the potential hazards of synthetic preservatives has led to a growing demand for natural alternatives. This study was designed to evaluate the antimicrobial potential of postbiotics derived from lactiplantibacillus plantarum and Lacticaseibacillus casei against common spoilage and pathogenic microorganisms in cheese, as well as to investigate their impact on the microbiological and chemical properties of cheese during storage.
Material and Methods: Postbiotics were extracted from cultures of L. plantarum and L. casei and their antimicrobial activities were tested using standard microbiological assays against Gram-positive and Gram-negative bacteria. To assess practical application, the postbiotics were applied as coatings on cheese samples either alone or in combination with whey protein concentrate (WPC). Microbiological counts, chemical parameters, and sensory evaluation were performed throughout storage. Data were analyzed to determine the comparative effectiveness of treatments.
Results and Conclusion: The findings showed that the postbiotic derived from L. plantarum demonstrated stronger antimicrobial effects, particularly against Gram-positive bacteria, compared to that from L. casei. However, when combined with WPC, the antimicrobial activity of both postbiotics declined. Despite this limitation, postbiotics applied alone significantly reduced microbial counts during storage without altering the main chemical properties of the cheese. Sensory evaluation confirmed the overall acceptability of postbiotic and postbiotic-WPC treated samples. In conclusion, postbiotics can serve as promising natural antimicrobial agents in cheese preservation, though further optimization is required to enhance their activity when combined with protein-based carriers such as WPC.
Keywords: Antimicrobial activity, Cell-free supernatant, Cheese, Lactic acid bacteria, Postbiotic
- Introduction
The food industry consistently faces substantial challenges from pathogenic and spoilage microorganisms, which are major drivers of foodborne diseases (FBDs), product quality degradation, and significant economic losses [1]. Globally, FBDs remain a pressing public health issue, with more than 600 million cases and nearly 420,000 deaths reported annually due to contaminated food and water [2,3]. These statistics have reinforced the urgency of strengthening food safety measures and minimizing contamination throughout production, processing, and storage stages [4,5]. At the same time, consumer concerns over the potential health risks associated with chemical preservatives, alongside the increasing demand for minimally processed foods, have fueled interest in natural preservation strategies [6,7]. Such approaches not only lessen dependence on artificial additives but also align with the growing trend toward clean-label products.
In this regard, biological protection has emerged as a promising strategy, harnessing beneficial microorganisms and their antimicrobial metabolites to suppress the proliferation of spoilage and pathogenic organisms [4]. Probiotics, especially lactic acid bacteria (LAB), have drawn considerable attention for their ability to generate bioactive substances such as organic acids, hydrogen peroxide, diacetyl, and bacteriocins [8]. Among these, LAB species such as Lactobacillus and Bifidobacterium are generally recognized as safe (GRAS) and have been widely employed for decades in the fermentation of diverse foods, including dairy products, cereals, and vegetables [9]. More recently, growing attention has been directed toward postbiotics, defined as non-viable microbial cells, cellular components, or metabolites that provide functional health benefits [10]. Compared with probiotics, postbiotics present notable practical advantages: they are independent of cell viability, exhibit greater stability under processing and storage conditions, and pose a lower risk of antibiotic resistance or incompatibility with food matrices. Additionally, postbiotics retain antimicrobial activity across a wide range of pH and temperature conditions, are capable of disrupting pathogenic biofilms, and can neutralize harmful contaminants such as pesticides and mycotoxins. In food systems, they have been investigated both as direct additives and as functional components of active packaging technologies, thereby overcoming limitations associated with the use of live microbial cultures [4].
Cheese provides a particularly critical application, as it is highly vulnerable to microbial contamination during both processing and storage. Among potential threats, Listeria monocytogenes is of significant concern due to its ability to withstand stress conditions and its high fatality rate in human infections [11,12]. Epidemiological evidence has repeatedly linked outbreaks of listeriosis to cheeses manufactured from raw or inadequately pasteurized milk [13], underscoring the urgent demand for innovative, safe, and effective antimicrobial approaches in dairy preservation.
Recent investigations have demonstrated that postbiotics and bacteriocin-like compounds derived from lactic acid bacteria (LAB) are capable of inhibiting pathogenic bacteria across diverse food matrices, including meat, seafood, and dairy products [14–19].
In parallel, whey protein, a major by-product of cheese production, has attracted considerable interest as a functional material for edible coatings and packaging, owing to its excellent barrier properties and strong film-forming capacity [11]. Incorporating antimicrobial postbiotics into whey protein systems may therefore offer dual advantages: enhancing microbial safety and prolonging shelf life while preserving desirable sensory characteristics.
Although evidence supporting postbiotics as natural preservatives is growing, relatively few studies have systematically assessed their performance in real cheese systems, particularly when combined with whey protein–based coatings. This gap in knowledge constrains a comprehensive understanding of their effectiveness under practical conditions.
The present study aims to bridge this gap by examining the antimicrobial activity of postbiotics derived from lactiplantibacillus plantarum and Lacticaseibacillus casei against key dairy pathogens, while also evaluating the microbial and chemical quality of cheese coated with whey protein concentrate (WPC) enriched with these postbiotics. This integrated strategy offers a novel, effective, and sustainable approach to improving food safety and preservation in dairy products.
- Materials and Methods
2.1 Study design
This descriptive-analytical study was conducted between 22 November 2022 and 15 September 2023. The study protocol was reviewed and approved by the Medical Ethics Committee of Gonabad University of Medical Sciences (IR.GMU.REC.1401.073).
2.2 Bacterial strains
Three lactiplantibacillus plantarum strains were isolated from traditional Iranian cheeses [20]. In addition, one Lacticaseibacillus casei strain (1608 PTCC, IBRC of Iran), Listeria monocytogenes (7644 ATCC), Escherichia coli (1338 PTCC), and Staphylococcus aureus (1431 PTCC) were obtained from the Laboratory of Specialization in Nutrition, Mashhad University of Medical Sciences.
2.3 Postbiotic preparation
Each lactic acid bacterial strain was cultured separately in MRS broth medium and incubated under anaerobic conditions at 37 °C for 24 hours. The cultures were then centrifuged at 6000 rpm for 10 minutes at 4 °C. The resulting cell-free supernatants were filtered through a 0.4 µm membrane filter and subsequently freeze-dried (freezing temperature −83 °C, pump pressure 0.0026 mBar, storage temperature −60 °C) for use in subsequent experiments [21].
2.4 Chemical analysis of cell-free supernatants of Lactobacillus sp.
The chemical compounds of postbiotics were identified following the method described by Ryan et al. (2009), with minor modifications. For derivatization, 1 mL of the supernatant was mixed with 10 mL of absolute ethanol and 15 drops of sulfuric acid (97%), and the mixture was stirred at 80 °C for one hour. After cooling, 20 mL of distilled water was added, and extraction was performed five times with 50 mL of dichloromethane, collecting the lower phase each time. The pooled extracts were combined with 50 g of sodium sulfate and passed through filter paper. The solvent was then removed using a vacuum evaporator at 50 °C, and the remaining residue was injected into the gas chromatography–mass spectrometry (GC–MS) system.
The chemical composition of the derivatized postbiotics was analyzed using a GC instrument (Agilent HP-6890, Agilent Technologies, Palo Alto, CA, USA) operated with Agilent GC/MS Mass Hunter Acquisition software. The GC system was equipped with an Agilent HP-5ms column (30 m length, 0.25 mm inner diameter, 0.25 μm film thickness). Helium was used as the carrier gas at a flow rate of 1 mL/min. The oven temperature was programmed to increase from 110 °C to 240 °C at a rate of 4 °C/min with no hold time. A 10 μL sample was injected with a 5:1 split ratio [22].
2.5 Antimicrobial Activity In-vitro
2.5.1 Agar-well diffusion
Three pathogens (L. monocytogenes, E. coli, and S. aureus) were inoculated separately at a concentration of 5 log10 CFU/mL onto the surface of Muller Hinton Agar (MHA) plates. Wells with an 8 mm diameter were then created in the agar, and 100 µL of postbiotic suspensions at concentrations of 5%, 10%, and 20% were added to the wells. Plates were incubated aerobically at 37 °C for 24–48 hours. Nisin (625 IU/mL) and sterile distilled water served as the positive and negative controls, respectively. Antimicrobial activity was expressed as the mean diameter (mm) of inhibition zones, measured as the clear areas surrounding the wells. Each assay was performed in triplicate [23].
2.5.2 Determination of minimum inhibitory concentra-tion (MIC) and minimum bactericidal concentration (MBC) by the microdilution method
MIC and MBC values were determined according to the Clinical and Laboratory Standards Institute (CLSI, 2017) guidelines. After 24 hours of aerobic incubation at 37 °C, the wells were examined for turbidity. To determine the MBC, 10 µL samples from wells without turbidity (corresponding to MIC and higher concentrations) were streaked onto MHA plates in triplicate and incubated under the same conditions [24].
2.5.3 Anti-listeria activity of coatings containing postbiotics
To prepare the coating solution, 50 mL of deionized water was heated to 90 °C in a bain-marie, and 4.34 g of whey protein concentrate (WPC; protein 81.2%, lactose 7.4%, fat 6%, moisture 5%, ash 4%, pH 6.1; Alinda, Greece) was fully dissolved. The solution was maintained at this temperature for 45–60 minutes, during which 2.71 g of glycerol and 0.081 g of Tween 80 were added. After cooling, cell-free supernatants (CFS) were incorporated at concentrations corresponding to MIC and MBC [25]. The anti-listeria activity of coatings containing postbiotics was then evaluated using the agar-well diffusion method. CFS without coating, at equivalent concentrations, was included as a positive control [26].
2.6 Inoculation and coating of cheese samples
A pasteurized traditional cheese, commercially available in local markets, was selected for this study. The cheeses analyzed were pasteurized varieties inoculated with a fungal starter culture. Four treatment groups were prepared to evaluate microbial and chemical characteristics over storage on days 0, 1, 2, 4, 6, 8, and 10 (Table 1).
Cheese pieces of approximately 10 g (3 × 3 × 1 cm) were inoculated with L. monocytogenes at a level of 1 × 10⁵ CFU/g by spreading 1 mL of an appropriately diluted suspension onto the surface [18]. Samples were then allowed to stabilize to ensure bacterial adherence. The treatments were as follows:
- Inoculated cheese without further treatment,
- Cheese immersed in CFS solution at the designated concentration,
- Cheese coated with WPC solution without CFS, and
- Cheese coated with WPC solution containing CFS (immersion for 4–5 minutes).
All samples were stored at 4 °C until further analysis.
2.7 Microbial analyses in-situ
- monocytogenes, total viable microorganisms, molds, and yeasts were enumerated using Palcam Agar, PCA, and SDA media, respectively. Palcam and PCA plates were incubated at 37 °C for 48 hours, while SDA plates were incubated at 25 °C for 3–5 days [26]. The enumeration of molds and yeasts was carried out according to the Iranian National Standard No. 2406: Microbiology of milk and milk products — Specifications and test methods [27].
2.8 Chemical analyses
The pH of cheese samples was measured using a pre-calibrated pH meter, and moisture content was determined by the gravimetric method on the designated sampling days [17,21].
2.9 Sensory analyses
Cheese slices coated with cell-free supernatants, free of L. monocytogenes, were evaluated for taste, color, aroma, texture, and overall acceptability by a panel of 10 semi-trained assessors using a 5-point hedonic scale [15]. All participants were adults above the legal age and voluntarily provided written informed consent in compliance with ethical standards for human subject research. Evaluations were conducted under identical environmental and temporal conditions to ensure consistency.
2.10 Statistical analyses
Statistical analyses were performed using SPSS software version 26. Mean values from three independent replicates were compared between two groups using the independent t-test, while comparisons among more than two independent groups were carried out using one-way ANOVA. Changes in data trends over the 10-day storage period were analyzed using one-way repeated measures ANOVA. A p-value of <0.05 was considered statistically significant.
- Results and Discussion
3.1 Antimicrobial activity of postbiotics
Postbiotics derived from both Lactobacillus species demonstrated inhibitory effects against the three tested pathogens, with higher postbiotic concentrations corresponding to stronger antimicrobial activity (Fig. 1).
Fig. 1. Antimicrobial activity of postbiotics against L. monocytogenes evaluated by the agar-well diffusion method.
The greatest inhibition was observed with the postbiotic from L. plantarum at a 20% concentration against L. monocytogenes, producing an inhibition zone of 30.67 ± 0.57 mm. In contrast, the weakest inhibition was observed with the postbiotic from L. casei at a 5% concentration against S. aureus, yielding an inhibition zone of 8.63 ± 0.55 mm. Across all concentrations, the postbiotic of L. plantarum exhibited significantly stronger inhibitory activity against L. monocytogenes and S. aureus compared to that of L. casei (p < 0.05) (Tables 2–4). For E. coli, no significant differences were detected between the two postbiotics except at the 10% concentration (Table 3). These findings suggest that postbiotics from L. plantarum are more effective against Gram-positive pathogens than those from L. casei.
Overall, the results indicate that L. plantarum postbiotics exert stronger antimicrobial effects against Gram-positive bacteria, with the most consistent reductions achieved through the “CFS only” treatment rather than the CFS–WPC combination. The maximum reduction compared to control (~0.80 log CFU/g for L. monocytogenes at day 2) declined over subsequent storage days, while pH, moisture, and sensory acceptability remained unaffected. This strain- and target-dependent pattern is consistent with previous evidence showing that LAB-derived cell-free supernatants inhibit Gram-positive pathogens primarily through organic acids and bacteriocin-like metabolites, mechanisms that involve pH reduction and disruption of microbial membranes [4,6,9,28–30]. Arena et al. also reported strong anti-pathogen activity of L. plantarum supernatants, with acidification identified as a major contributing factor [31,32]. The reduced inhibitory effect observed when CFS was incorporated into a whey protein carrier is consistent with the well-documented “matrix effects” described in the literature. Previous studies have shown that interactions between proteins and bioactive metabolites, along with the barrier properties of protein films, can delay the release and reduce the bioavailability of antimicrobial compounds [11,26,33–36]. Similar patterns of initial but transient inhibition, followed by partial recovery of pathogen populations, have been reported in fresh cheese, meat, and fish products treated with CFS- or bacteriocin-based films [15–17,19].
In agreement with these findings, the present study demonstrated that chemical attributes (pH and moisture) and sensory acceptance were not adversely affected, supporting the feasibility of integrating postbiotics into dairy preservation systems. However, further optimization of carrier composition and release kinetics is required to maximize antimicrobial effectiveness [37].
The MIC and MBC values of L. plantarum postbiotics against the three tested pathogens were determined as 31.25 mg/mL and 62.5 mg/mL, respectively, for Gram-positive bacteria, and 125 mg/mL for E. coli. These results highlight a greater inhibitory effect against Gram-positive bacteria at lower concentrations (Table 5).
The postbiotic of L. casei exhibited a comparable inhibitory effect against L. monocytogenes to that of L. plantarum, except at the 5% concentration, where a difference was observed in comparison with E. coli (Table 6). No significant differences were detected in the inhibition of S. aureus and E. coli (p > 0.05) (Table 7).
The MIC of L. casei postbiotics against L. monocytogenes was higher (62.5 mg/mL) than that observed for L. plantarum, although both species showed identical MBC values. For E. coli, the MBC of L. casei postbiotics was lower than that of L. plantarum. In contrast, for S. aureus, the MIC and MBC values were the same for both postbiotics.
Arrioja et al. (2020) similarly reported that CFS derived from L. plantarum exhibited stronger inhibitory activity against most pathogens compared with CFS from L. casei [33]. Arena et al. (2016) further demonstrated variability in inhibition zones and MICs among different L. plantarum strains against various pathogens, with generally greater effects observed against Gram-positive bacteria [31]. Consistent with the present findings, Tenea and Barrigas (2018) showed that bacteriocin-containing supernatants from L. plantarum (Cys5-4) exhibited variable inhibitory effects against two E. coli strains [7]. Koohestani et al. (2018) also reported that CFS from L. casei 431 produced an inhibition zone of 13 mm against S. aureus, which closely aligns with the current results [23]. In contrast, Yordshahi et al. (2020) documented smaller inhibition zones for L. plantarum postbiotics against L. monocyte-genes compared with those observed in this study [21].
The antimicrobial activity of lactic acid bacteria has been attributed to a range of metabolites, including organic acids, polyamines, proteases, and bacteriocins. The effectiveness of postbiotics depends on multiple factors such as bacterial strain, metabolite composition and concentration, preparation method, and pathogen type. Numerous studies have confirmed that Gram-positive bacteria are generally more susceptible to the antagonistic compounds in postbiotics than Gram-negative bacteria, consistent with the findings of the present work [38].
Results from the agar-well diffusion assays indicated that postbiotics incorporated into WPC coating solutions at MBC concentrations generally exhibited reduced inhibition against L. monocytogenes compared to postbiotics applied alone (Table 8). However, the combination of L. plantarum postbiotics with WPC coatings produced greater inhibition than L. casei postbiotics at equivalent concentrations.
Based on the in-vitro assays, the L. plantarum postbiotic at twice the MBC concentration was identified as the most effective formulation and was subsequently selected for testing in the food model.
3.2 Identification of chemical compounds of extracted CFS
The chemical compounds identified in the postbiotics derived from L. plantarum and L. casei are presented in their respective chromatograms (Figs. 2 and 3).
Sezen Özcelik et al. (2016) reported that LAB strains are particularly efficient producers of succinic acid, especially when cultivated in MRS broth. Succinic acid serves as a key intermediate in the Krebs cycle and a common fermentation byproduct, reflecting the strong metabolic capacity of LABs. The quantity and composition of organic acids produced by LABs vary considerably across strains and culture media, with pH and temperature exerting significant influence [28]. Similarly, Iqbal Hossain et al. (2021) identified nine distinct organic acids, including succinic acid, in several LAB strains such as L. plantarum, highlighting the diverse metabolite production potential of these bacteria [29]. In the present study, the derivatization technique applied proved particularly effective in detecting compounds such as esters and alkanes.
Shehata et al. demonstrated that LABs synthesize antifungal metabolites that differ across strains, with organic acids and hydrogen peroxide serving as the primary contributors to antifungal activity. Their study identified compounds such as pentadecane and 2,4-di-tert-butylphenol, both known to inhibit foodborne pathogens and fungi. Additionally, antimicrobial compounds including 6-octadecenoic acid methyl ester and hexadecanoic acid methyl ester—also detected in the current work—were previously reported. Deepthi et al. further showed that LABs generate a wide variety of antifungal carboxylic acid esters, whose effectiveness depends on strain variability and fatty acid chain length. For example, 10-octadecenoic acid methyl ester, an unsaturated fatty acid ester with 19 carbon atoms, has been described as an effective antifungal metabolite. In agreement with Shehata et al., our findings also identified nonadecane, a compound without known antimicrobial activity [39,40].
Benzoic acid was detected in small quantities in both postbiotics. This compound, obtained from L. plantarum, has previously been reported by Siedler et al. as possessing antimicrobial properties [41].
In addition, several compounds not previously described for their antimicrobial effects were identified in the chemical analysis of the postbiotics, including phthalic acid, eicosane, chloroacetic acid, benzene propanoic acid, propenoic acid, triethyl citrate, isopropyl myristate, butanedioic acid, and tetradecanoic acid.
3.3 Microbiological analyses
The growth of L. monocytogenes was monitored in the control samples (treatment one), which demonstrated a continuous increase throughout 10 days of storage at 4 °C, with an overall rise of 1.1 log CFU/g (Fig. 4).
Fig. 4. Growth of L. monocytogenes across four food treatments during 10 days of storage at 4 °C.
A significant reduction in pathogen counts was observed in the second treatment, which contained CFS, compared with the other three treatments over the 10-day storage period (p < 0.05). In contrast, no significant differences were detected in contamination levels between the third and fourth treatments and the control (p > 0.05). The largest reduction in L. monocytogenes relative to the control was 0.80 log CFU/g, recorded in the second treatment on day two; however, this difference gradually declined over the storage period (Table 9).
The results for total microbial counts (Fig. 5) and mold and yeast counts (Fig. 6) mirrored those observed for L. monocytogenes, with only the second treatment showing a significant reduction (p < 0.05) compared with the other treatments over the 10-day storage period. Fig. 5. Total microbial counts across four food treatments during 10 days of storage at 4 °C. Fig. 6. Mold and yeast count across four food treatments during 10 days of storage at 4 °C.
These findings are consistent with the study by Iqbal Hossain et al. (2021), which reported differences in the inhibitory effect (MEC) of CFS derived from L. plantarum when tested in a culture medium (TSB) compared with a food model (whole milk) against L. monocytogenes [29]. Similarly, Hartmann et al. (2011) demonstrated that the efficiency of bacterial fermentations varied depending on the food matrix, with bacteriocin IDE0105 from L. plantarum showing weaker activity in milk than in BHI medium. Such discrepancies may be explained by limited solubility or uneven distribution of antimicrobial compounds in food systems, inactivation of antagonistic chemicals by food components or microflora-derived enzymes, or interactions between antimicrobials and food ingredients [34].
Comparable to the present study, Jutinico-Shubach et al. (2020) reported a 0.83 log CFU/g reduction in L. monocytogenes in cheese samples treated with CFS from Pediococcus pentosaceus 147 on day two of storage compared with the control. This closely aligns with the 0.80 log CFU/g reduction observed in our second treatment at the same time point [17]. Similarly, Salvucci et al. (2019) found reductions in L. monocytogenes in cheese treated with films containing bacteriocins from Enterococcus faecium ES216 [18]. Marques et al. (2017) also observed that L. monocytogenes counts in actively packaged cheese samples treated with CFS from Latilactobacillus curvatus P99 reached levels comparable to the control by day 10. Moreover, films containing CFS at MIC concentrations (15.6 µL/mL) did not significantly reduce bacterial counts compared with controls, consistent with our findings [26]. Yordshahi et al. (2020) likewise reported the survival of L. monocytogenes in all meat samples treated or untreated with active packaging containing postbiotics from L. plantarum (ATCC 14917), with a 1.0 log CFU/g increase in control samples by day nine—similar to the 1.1 log CFU/g increase observed in our study [21].
The type of film or coating, as well as the nature of the food, plays a critical role in the release of antimicrobial agents from polymer matrices into food systems [21]. Beristain-Bauza et al. (2017) demonstrated that untreated beef samples exhibited a modest reduction (<0.5 log CFU/g) in L. monocytogenes Scott A growth after five days, whereas beef treated with WPI films containing CFS from Latilactobacillus sakei (NRRL B-1917) showed a reduction of 1.4 log CFU/g after the same period [19]. Such variability highlights the dependence of antimicrobial efficacy on factors including film or coating type, bacterial strain, food model, and release kinetics of antagonistic compounds.
The study by Beristain-Bauza et al. (2016) suggested that the stronger antimicrobial performance of WPI films may be attributed to the hydrophobic nature of whey proteins, which facilitates compatibility with hydrophilic CFS obtained from Lacticaseibacillus rhamnosus (NRRL B-442). This mechanism may help explain the diminished inhibitory effect observed in the present study when CFS was combined with WPC coatings [38].
3.4 Chemical analyses
The pH values of cheese samples from the different treatments showed no significant differences (p > 0.05), with levels ranging from 6.50 to 6.56. This observation is consistent with Jutinico-Shubach et al. (2020), who reported comparable pH values in both control and CFS-treated cheese samples derived from Pediococcus pentosaceus 147 [17]. Similarly, Salvucci et al. (2019) found no significant differences in moisture content between control and treated cheese samples during storage, further corroborating the findings of the present study [18].
3.5 Sensory analyses
Sensory evaluation revealed no significant differences (p > 0.05) in overall acceptability or odor index between the control treatment and the WPC–CFS combination treatment (Fig. 7).
No significant differences were detected between the two CFS treatments and the WPC–CFS combination. However, a significant difference (p < 0.05) was observed in the color parameter of the control treatment compared with the other treatments, likely attributable to the MRS culture medium used as the primary substrate for LAB growth and CFS production (Table 10). This finding aligns with Beristain-Bauza et al. (2016), who reported pronounced color differences in WPI films containing CFS from Lacticaseibacillus rhamnosus (NRRL B-442) [37].
In contrast, no significant differences were detected in texture or taste across treatments (p > 0.05), indicating good sensory acceptance of the treated samples. These results are in line with those of Beristain-Bauza et al. (2017), who observed favorable sensory evaluations in grilled beef treated with WPI films containing CFS from Latilactobacillus sakei (NRRL B-1917) [19].
- Conclusion
This study demonstrated that postbiotics derived from L. plantarum and L. casei possess significant antimicrobial activity against major foodborne pathogens when applied as coatings in pasteurized cheese. Among the two, L. plantarum exhibited stronger inhibitory effects, particularly against Gram-positive bacteria. However, incorporation of postbiotics into WPC-based coatings reduced their antimicrobial efficacy, likely due to matrix interactions. Importantly, postbiotic treatments successfully suppressed microbial growth during refrigerated storage without affecting the key chemical characteristics of cheese and were well accepted in sensory evaluations.
These findings underscore the potential of postbiotic-based coatings as natural, consumer-friendly alternatives to chemical preservatives in dairy products. Beyond cheese, this strategy could be extended to a broad range of perishable foods where safety, shelf life, and sensory acceptance are of critical importance. Nevertheless, further research is required to optimize carrier systems, enhance stability, and evaluate large-scale industrial applications across diverse food matrices.
- Acknowledgements
The authors would like to thank Mashhad University of Medical Sciences, Mashhad, Iran, and Gonabad University of Medical Sciences, Gonabad, Iran for their support.
- Declaration of competing interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
- Authors’ Contributions
Zeinab Hadadfar: Conceptualization; Data curation; Formal analysis; Investigation; Methodology; Software; Supervision; Visualization; Writing – original draft. Asma Afshari: Conceptualization; Investigation; Methodology; Resources; Supervision; Validation; Writing – review and editing. Alireza Mohammadzadeh: Investigation; Methodology; Resources; Supervision; Validation; Writing – review and editing. Zohreh Abdimoghadam: Funding acquisition; Investigation; Project administration; Resources; Supervision; Validation; Writing – review and editing.
Funding: This work was supported by the Gonabad University of Medical Sciences [grant numbers 1130].
- Using Artificial Intelligent Chatbots
The authors declare no artificial intelligent chatbot use.
- Ethical Consideration
The study protocol was reviewed and approved by the Medical Ethics Committee of Gonabad University of Medical Sciences (IR.GMU.REC.1401.073).
- Antimicrobial activity
- Cell-free supernatant
- Cheese
- Lactic acid bacteria
- Postbiotic
ارجاع به مقاله
مراجع
Tauxe RV, Doyle MP, Kuchenmüller T, Schlundt J, Stein C. Evolving public health approaches to the global challenge of foodborne infections. Int J Food Microbiol. 2010; 139 Suppl 1: S16-28. https://doi.org/10.1016/j.ijfoodmicro.2009.10.014
Singh N, Kaur R, Singh BP, Rokana N, Goel G, Puniya AK, et al. Impairment of Cronobacter sakazakii and Listeria monocytogenes biofilms by cell-free preparations of lactobacilli of goat milk origin. Folia Microbiol. 2020; 65(1):185-196. https://doi.org/10.1007/s12223-019-00708-3
Hashemi M, Asadi Touranlou F, Adibi S, Afshari A, Shakeri G. Foodborne bacteria in Iran: A 23-year systematic review of high-risk foods. Iran J Vet Sci Technol. 2024; 16(1):15-28. https://doi.org/10.22067/ijvst.2024.87069.1357
Moradi M, Kousheh SA, Almasi H, Alizadeh A, Guimarães JT, Yılmaz N, et al. Postbiotics produced by lactic acid bacteria: The next frontier in food safety. Compr Rev Food Sci Food Saf. 2020; 19(6): 3390-3415. https://doi.org/10.1111/1541-4337.12609
Patrovský M, Kouřimská L, Havlíková Š, Marková J, Pechar R, Rada V. Utilization of bacteriocin-producing bacteria in dairy products. Mljekarstvo. 2016; 66(3): 215-224. https://doi.org/10.15567/mljekarstvo.2016.0306
Silva CC, Silva SP, Ribeiro SC. Application of bacteriocins and protective cultures in dairy food preservation. Front Microbiol. 2018; 9: 594. https://doi.org/10.3389/fmicb.2018.00594
Tenea G, Barrigas A. The efficacy of bacteriocin-containing cell-free supernatant from Lactobacillus plantarum Cys5-4 to control pathogenic bacteria growth in artisanal beverages. Int Food Res J. 2018; 25(5): 2031-2037.
Moghimani M, Onyeaka H, Hashemi M, Afshari A. Evaluation of the probiotic microorganisms isolated from Iranian milk kefir beverages. Front Microbiol. 2024; 15: 1385301. https://doi.org/10.3389/fmicb.2024.1385301
Mani-López E, Arrioja-Bretón D, López-Malo A. The impacts of antimicrobial and antifungal activity of cell-free supernatants from lactic acid bacteria in vitro and foods. Compr Rev Food Sci Food Saf. 2022; 21(1): 604-641. https://doi.org/10.1111/1541-4337.12838
Homayouni-Rad A, Fathi-Zavoshti H, Douroud N, Shahbazi N, Abbasi A. Evaluating the role of postbiotics as a new generation of probiotics in health and diseases. J Ardabil Univ Med Sci. 2020; 19(4): 381-399. https://doi.org/10.29252/jarums.19.4.381
Ramos OL, Fernandes JC, Silva SI, Pintado ME, Malcata FX. Edible films and coatings from whey proteins: A review on formulation, and on mechanical and bioactive properties. Crit Rev Food Sci Nutr. 2012; 52(6): 533-552. https://doi.org/10.1080/10408398.2010.500528
Fox PF, Guinee TP, Cogan TM, McSweeney PLH. Fundamentals of Cheese Science. 2nd ed. New York: Springer; 2017. https://doi.org/10.1007/978-1-4899-7681-9
Centers for Disease Control and Prevention (CDC). Outbreaks of listeriosis in cheese and meats. Atlanta: CDC; 2019. available at: https://www.cdc.gov/listeria/index.html
İncili GK, Karatepe P, Akgöl M, Tekin A, Kanmaz H, Kaya B, et al. Impact of chitosan embedded with postbiotics from Pediococcus acidilactici against emerging foodborne pathogens in vacuum-packaged frankfurters during refrigerated storage. Meat Sci. 2022; 188: 108786. https://doi.org/10.1016/j.meatsci.2022.108786
Jo DM, Park SK, Khan F, Kang MG, Lee JH, Kim YM. An approach to extend the shelf life of ribbonfish fillet using lactic acid bacteria cell-free culture supernatant. Food Control. 2021; 123: 107731. https://doi.org/10.1016/j.foodcont.2020.107731
da Costa RJ, da Silva AP, da Fonseca RN, de Oliveira Hübner S, Nalério ES, de Lima Marques J, et al. Characterization of Enterococcus faecium EO1 and its bacteriocin-like inhibitory substance for control of Listeria monocytogenes in fresh mutton sausage. LWT. 2021; 141: 110954. https://doi.org/10.1016/j.lwt.2021.110954
Jutinico-Shubach A, Gutiérrez-Cortés C, Suarez H. Antilisterial activity of chitosan-based edible coating incorporating cell-free supernatant from Pediococcus pentosaceus 147 on the preservation of fresh cheese. J Food Process Preserv. 2020; 44(9): e14715. https://doi.org/10.1111/jfpp.14715
Salvucci E, Rossi M, Colombo A, Pérez G, Borneo R, Aguirre A. Triticale flour films added with bacteriocin-like substance (BLIS) for active food packaging applications. Food Packag Shelf Life. 2019; 19:193-199. https://doi.org/10.1016/j.fpsl.2018.05.007
Beristain-Bauza SC, Mani-López E, Palou E, López-Malo A. Antimicrobial activity of whey protein films supplemented with Lactobacillus sakei cell-free supernatant on fresh beef. Food Microbiol. 2017; 62: 207-211. https://doi.org/10.1016/j.fm.2016.10.024
Afshari A, Hashemi M, Tavassoli M, Eraghi V, Noori SMA. Probiotic bacteria from 10 different traditional Iranian cheeses: Isolation, characterization, and investigation of probiotic potential. Food Sci Nutr. 2022; 10(6): 2009-2020. https://doi.org/10.1002/fsn3.2817
Yordshahi AS, Moradi M, Tajik H, Molaei R. Design and preparation of antimicrobial meat wrapping nanopaper with bacterial cellulose and postbiotics of lactic acid bacteria. Int J Food Microbiol. 2020; 321: 108561. https://doi.org/10.1016/j.ijfoodmicro.2020.108561
Ryan LAM, Bello FD, Czerny M, Koehler P, Arendt EK. Quantification of phenyllactic acid in wheat sourdough using high resolution gas chromatography–mass spectrometry. J Agric Food Chem. 2009; 57(3):1060-1064. https://doi.org/10.1021/jf802578e
Koohestani M, Moradi M, Tajik H, Badali A. Effects of cell-free supernatant of Lactobacillus acidophilus LA5 and Lactobacillus casei 431 against planktonic form and biofilm of Staphylococcus aureus. Vet Res Forum. 2018; 9(4): 301-306. https://doi.org/10.30466/vrf.2018.33086
Jalilzadeh A, Hesari J, Peighambardoust SH, Javidipour I. The effect of whey protein-based edible coating containing natamycin and lysozyme-xanthan gum conjugate on the shelf life of ultrafiltrated white cheese. J Food Bioproc Eng. 2020; 3(2): 168-177. https://doi.org/10.22059/jfabe.2020.306083.1068
de Lima Marques J, Funck GD, da Silva Dannenberg G, dos Santos Cruxen CE, El Halal SLM, Dias ARG, et al. Bacteriocin-like substances of Lactobacillus curvatus P99: Characterization and application in biodegradable films for control of Listeria monocytogenes in cheese. Food Microbiol. 2017; 63:159-163. https://doi.org/10.1016/j.fm.2016.11.008
Iran National Standards Organization (INSO). Microbiology of milk and milk products-Specifications and test methods. 4th revision. National Standard No. 2406. Tehran: INSO; 2024.
Özcelik S, Kuley E, Özogul F. Formation of lactic, acetic, succinic, propionic, formic and butyric acid by lactic acid bacteria. LWT. 2016; 73:536-542. https://doi.org/10.1016/j.lwt.2016.05.067
Hossain MI, Kim K, Mizan MFR, Toushik SH, Ashrafudoulla M, Roy PK, et al. Comprehensive molecular, probiotic, and quorum-sensing characterization of anti-listerial lactic acid bacteria, and application as bioprotective in a food (milk) model. J Dairy Sci. 2021; 104(6): 6516-6534. https://doi.org/10.3168/jds.2020-19034
Zapaśnik A, Krakowska-Sobocka Z, Radecka-Poltone R, et al. Role of lactic acid bacteria in food preservation and safety. Foods. 2022; 11(9): 1283. https://doi.org/10.3390/foods11091283
Arena MP, Silvain A, Normanno G, Grieco F, Drider D, Spano G, et al. Use of Lactobacillus plantarum strains as a bio-control strategy against food-borne pathogenic microorganisms. Front Microbiol. 2016; 7: 464. https://doi.org/10.3389/fmicb.2016.00464
Isaac-Bamgboye FJ, Adewale OB, Komolafe TR, et al. Exploring the potential of postbiotics for food safety and human health improvement. J Nutr Metab. 2024; 2024: 1868161. https://doi.org/10.1155/2024/1868161
Arrioja-Bretón D, Mani-López E, Palou E, López-Malo A. Antimicrobial activity and storage stability of cell-free supernatants from lactic acid bacteria and their applications with fresh beef. Food Control. 2020; 115: 107286. https://doi.org/10.1016/j.foodcont.2020.107286
Hartmann HA, Wilke T, Erdmann R. Efficacy of bacteriocin-containing cell-free culture supernatants from lactic acid bacteria to control Listeria monocytogenes in food. Int J Food Microbiol. 2011; 146(2): 192–199. https://doi.org/10.1016/j.ijfoodmicro.2011.02.031
Bangar SP, Teixeira JA, Silva CCG. Recent advances in the use of edible films and coatings with probiotic and bacteriocin-producing lactic acid bacteria. Food Biosci. 2023; 56: 103196. https://doi.org/10.1016/j.fbio.2023.103196
Silva SPM, Teixeira JA. Application of enterocin-whey films to reduce Listeria monocytogenes contamination on ripened cheese. Food Microbiol. 2023; 109: 104134. https://doi.org/10.1016/j.fm.2022.104134
Dong J, Wang S, Li M, et al. Application of a chitosan-based active packaging film prepared with cell-free supernatant of Lacticaseibacillus paracasei ALAC-4 in Mongolian cheese preservation. J Food Prot. 2023; 86(12): 100158. https://doi.org/10.1016/j.jfp.2023.100158
Beristain-Bauza SC, Mani-López E, Palou E, López-Malo A. Antimicrobial activity and physical properties of protein films added with cell-free supernatant of Lactobacillus rhamnosus. Food Control. 2016; 62: 44-51. https://doi.org/10.1016/j.foodcont.2015.10.010
Shehata MG, Badr AN, El Sohaimy SA, Asker D, Awad TS. Characterization of antifungal metabolites produced by novel lactic acid bacterium and their potential application as food biopreservatives. Ann Agric Sci. 2019; 64(1): 71-78. https://doi.org/10.1016/j.aoas.2019.05.002
Deepthi B, Poornachandra Rao K, Chennapa G, Naik M, Chandrashekara K, Sreenivasa M. Antifungal attributes of Lactobacillus plantarum MYS6 against fumonisin-producing Fusarium proliferatum associated with poultry feeds. PLoS One. 2016; 11(6): e0155122. https://doi.org/10.1371/journal.pone.0155122
Siedler S, Balti R, Neves AR. Bioprotective mechanisms of lactic acid bacteria against fungal spoilage of food. Curr Opin Biotechnol. 2019; 56:138-146. https://doi.org/10.1016/j.copbio.2018.11.015
- چکیده مشاهده شده: 307 بار
- pdf (English) دانلود شده: 15 بار