
Antidiabetic Bioactive Peptides from A2 Milk Fermented by Lacticaseibacillus rhamnosus: Enzyme Inhibition and Docking Insights
بیوتکنولوژی غذایی کاربردی,
دوره 12 شماره 1 (2025),
4 ژانویهٔ 2025
,
صفحه 1-15 (e21)
https://doi.org/10.22037/afb.v12i1.48611
چکیده
Background and Objective: Diabetes mellitus is a long-term disorder characterized by the body’s inability to regulate excessive blood glucose levels. The incidence of diabetes mellitus worldwide has significantly increased in recent decades. In addition to careful regulation of food quantities, improving food quality through the consumption of functional foods that do not trigger glucose spikes is also recommended. Accordingly, the aim of this research is to determine the potency of antidiabetic functional food derived from the fermentation of A2 cow's milk with higher β-casein content using Lacticaseibacillus rhamnosus RAL43.
Material and Methods: This study involved in vitro assays to test the inhibitory activity against diabetes-related enzymes, namely α-glucosidase, α-amylase, and dipeptidyl peptidase-4 (DPP4), followed by molecular docking simulations.
Results and Conclusion: The results of the study showed inhibitory activity against enzymes that trigger blood glucose spikes, namely 66.35% against α-glucosidase, 68.87% against α-amylase, and 10.69% against DPP4. The results of the analysis showed an increase in the quantity of peptides after fermentation, along with the results of the analysis of L. rhamnosus RAL43 which showed high proteolytic activity during fermentation. After ultrafiltration, it was found that the greatest inhibitory activity came from protein with molecular weight (MW) larger than 10 kDa. The amino acid sequencing process with high-resolution liquid chromatography-mass spectrometry then showed bioactive peptides, including VLVLDTDYK which was previously reported to show DPP4 inhibitory activity, along with many other peptides that display various specific bioactivities. The VLVLDTDYK peptide fragment was successfully docked and positioned in the DPP4 molecule through molecular docking simulations. This study concluded that A2 milk can be a functional substrate to produce specific bioactive peptides that inhibit enzymes that trigger blood glucose spikes that have a negative impact on diabetes. L. rhamnosus RAL43 can be developed as a proteolytic isolate and starter culture to produce foods with functional properties that can aid in blood glucose regulation.
Keywords: Bioactive Peptides, A2 Milk, Antidiabetic, Molecular Docking
Introduction
Diabetes mellitus (DM) has emerged as a significant global health issue. It is a persistent condition marked by increased blood glucose levels, typically above 180 mg.dL-1. Under normal circumstances, the hormone insulin regulates blood glucose levels by converting glucose into energy for cells. However, in individuals with diabetes, the body either does not produce enough insulin or cannot use it effectively to control blood glucose levels. According to data released by the International Diabetes Federation, Type 2 Diabetes Mellitus (T2DM) is the most common form globally and continues to increase significantly each year. The primary causes of T2DM include being overweight and obese, often due to an unbalanced diet and lack of physical activity. T2DM typically does not exhibit critical symptoms in its early stages, causing many individuals to remain unaware of their condition. Chatterjee et al. [1] and Antar et al. [2] identified a prediabetic phase that precedes full-blown diabetes. With proper prevention and management during the prediabetic phase, T2DM can still be avoided. In principle, individuals with T2DM must consistently avoid high blood glucose levels, particularly postprandial spikes [1,2]. Besides careful control of dietary quantity, improving diet quality through the consumption of functional foods that do not trigger glucose spikes is also recommended. Foods or beverages that offer health benefits are referred to as functional foods. Antidiabetic functional foods have been reported to originate from fermented products containing probiotics and/or bioactive compounds that inhibit digestive enzymes involved in carbohydrate breakdown into glucose, including α-amylase, α-glucosidase, and dipeptidyl peptidase-4 (DPP4) [3-5]. Functional foods with antidia-betic activity are considered a promising alternative for managing T2DM without the side effects often associated with pharmaceutical treatments [6].
Digestive enzyme inhibitors operate through various mechanisms. The enzymes α-amylase and α-glucosidase break down complex carbohydrates into simple sugars for absorption in the intestines. Inhibiting these enzymes helps slow glucose absorption and prevents postprandial blood glucose spikes [7]. Meanwhile, the DPP4 enzyme plays a critical role in blood glucose regulation by degrading incretin hormones such as GLP-1 (glucagon-like peptide-1) and GIP (glucose-dependent insulinotropic polypeptide). This degradation can suppress insulin secretion and burden the pancreas. DPP4 inhibitors are thought to prevent incretin breakdown, thereby maintaining normal glucose levels and enhancing insulin sensitivity [8]. Enzyme inhibitors targeting α-amylase, α-glucosidase, and DPP4 have been reported in the form of short-chain peptide fragments produced through proteolytic processes, either via enzy-matic hydrolysis or microbial fermentation [9-11]. These peptide-based inhibitors are thought to function as competitive inhibitors by attaching to the enzyme’s active site, preventing substrate binding, and thereby slowing down the buildup of glucose in the blood [12]. Other reported types of enzyme inhibitors include flavonoids, terpenoids, phenolic acids, tannins, alkaloids, and xanthones [10,13]. However, this study will focus specifically on exploring peptide-based inhibitors produced through fermentation by lactic acid bacteria (LAB) isolates.
Once the bioactivity data are obtained and the peptide sequences with potential inhibitory effects are identified, the next step is to perform molecular docking. Molecular docking is a crucial and efficient method for exploring the potential bioactivity of inhibitory peptides. It provides a strong scientific basis prior to biological testing, accelerates the screening process, and helps focus the research on the most promising peptide candidates [14]. This computational approach allows for the modeling and prediction of molec-ular interactions between candidate peptides and target enzymes. In this study, molecular docking is used to generate preliminary data supporting the hypothesis that the identified peptides may possess inhibitory activity against the target enzyme.
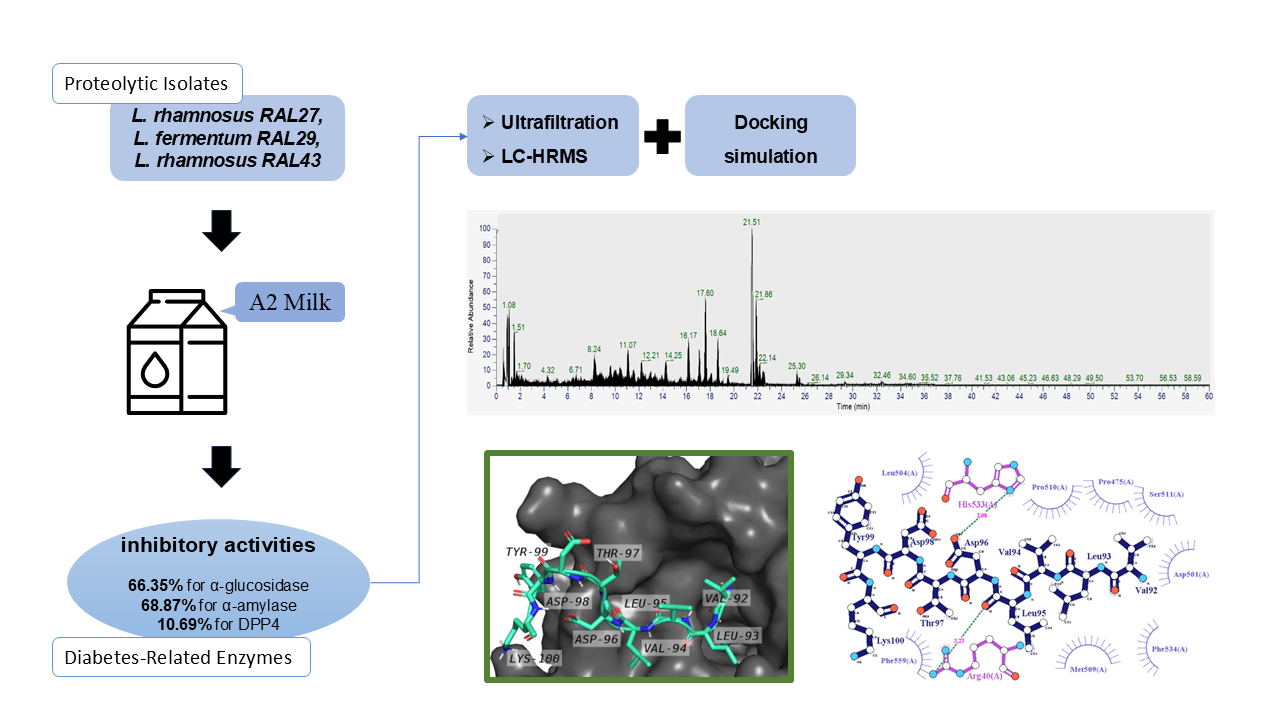
Fermented dairy products using LAB have been reported to exhibit antidiabetic potential [3,15,16]. Although not classified as therapeutic drugs, these products are considered effective as functional foods for diabetes preven-tion and management [7,17]. LAB have a significant advantage due to their proteolytic activity, enabling the breakdown of proteins into short-chain peptides. Dairy products fermented with LAB are well recognized as rich sources of bioactive peptides that exhibit antidiabetic, antioxidant, antihypertensive, cholesterol-lowering, and various other functional effects [18-20]. In this study, three lactic acid bacteria isolates (Lacticaseibacillus (L.) rhamnosus RAL27, Limosilactobacillus fermentum RAL29, and L. rhamnosus RAL43) previously isolated from Indonesian kefir grains [21], will be used to ferment A2-type cow’s milk. In recent years, A2 milk has been introduced. This type of milk was first commercialized in New Zealand and has since been marketed in several countries [22]. The main differences between A1 and A2 milk lie in the amino acid composition and β-casein. A1 β-casein contains histidine at position 67, while A2 β-casein contains proline at the same position [23]. These subtle differences can affect human digestion and metabolism. Some studies suggest that A1 milk may be associated with gastrointestinal discomfort and symptoms similar to lactose intolerance. A study by Choi et al. [24] found that participants consuming A1 milk reported more bloating and abdominal pain compared to those consuming A2 milk. The release of β-casomorphin-7 (BCM-7), a peptide derived from A1 β-casein during digestion, is thought to contribute to these symptoms. Some epidemiological studies suggest that A1 milk may be correlated with an increased risk of heart disease due to its inflammatory effects [23]. Conver-sely, A2 milk may offer cardioprotective effects, although further clinical trials are needed to validate these findings. A possible link between A1 milk consumption and the onset of type 1 diabetes has been proposed. A study by Kay et al. [23] suggests that early exposure to A1 milk may be associated with an increased risk of developing autoimmune conditions, possibly due to an immune response triggered by β-casomorphin-7 (BCM-7). With increasing health and wellness awareness, consumer demand for A2 milk has increased significantly. Many dairy companies have begun marketing A2 milk as a healthier alternative to A1 milk, capitalizing on its perceived benefits. This shift reflects a broader trend toward personalized nutrition and the importance of understanding genetic variation in food choices.The fermentation of A2 milk is expected to generate a new and unique peptide profile. Digestive conditions can influence the stability and absorption of peptides produced by A2 milk fermentation. Many peptides are biodegradable, so only peptides of a certain size or structure are retained. Smaller peptides may persist, while larger ones tend to break into fragments. Some bioactive peptides (e.g., DPP4 or α-glucosidase inhibitors) have been shown to remain active even after simulated in vitro digestion, but their persistence varies depending on sequence and structure [7,25]. However, not all peptides identified in vitro will persist into the systemic circulation. However, local activity in, for example, (α-glucosidase inhibition) is expected to remain relevant, even if the peptide is not absorbed in large quantities from the gut. Therefore, this study aims to determine the potential of antidiabetic functional food derived from the fermentation of A2 cow’s milk using lactic acid bacteria isolates, and to evaluate it through in vitro inhibitory activity tests against key diabetes-related enzymes: α-glucosidase, α-amylase, and DPP4, followed by binding simulation analysis.
- Materials and Methods
2.1. Material
This study used three isolates from Indonesian kefir grains: L. rhamnosus strain RAL27, Limosilactobacillus fermentum strain RAL29, and L. rhamnosus strain RAL43. The A2 milk (KIN Fresh Milk, Jakarta, Indonesia) and Mann Rogosa Sharpe broth (Himedia) were used as growth media. The chemicals used included α-glucosidase from Saccaromyces cerevisiae (Sigma-Aldrich Co.), α-amylase from from porcine pancreas (Sigma-Aldrich Co.), DPP4 inhibitor screening kit (Sigma-Aldrich Co.) included DPP4 enzyme from human, o-phthalaldehyde (Sigma-Aldrich Co.), p-Nitrophenyl α-D-glucopyranoside (PNPG; Sigma-Aldrich Co.), phenolphthalein (Himedia), Folin-Ciocalteu reagent (Sigma-Aldrich Co.), trichloroacetic acid (Himed-ia), methanol (Himedia), ethanol (Merck), sodium carbonate (Himedia), sodium pyrophosphate (Himedia), β-mercapto-ethanol (Merck), acarbose (Merck), Sitagliptin (Sigma-Aldrich Co.), phosphoric acid (Himedia), phosphate buffer saline (PBS; Himedia), 0.22µM filter membrane (Aijiren), and membrane filter 3.000 and 10.000 Da (Spin-X® UF Concentrators, Corning).
2.2 Isolates preparation
Isolates of L. rhamnosus strain RAL27, L. fermentum strain RAL29, and L. rhamnosus strain RAL43 were refreshed in Mann Rogosa Sharpe broth (MRSB) and incubated at 37oC for 48 h. Cultivation in MRSB was carried out 2 times and then the three isolates were adapted to A2 milk, by inoculating the isolates as much as 2% into A2 milk, then incubated at 37oC for 48 h. This process was also carried out 2 times.
2.3 Fermentation process
The fermentation process refers to the research of Yusuf et al. [26]. The isolate was inoculated to A2 milk at concen-tration of 2% from 5 log CFU.mL-1 starter. Incubation was conducted aerobically at 37°C for 24 h. The fermented milk obtained was characterized by assessing the total viability of LAB, pH value, acidity, aromatic profile, and coagulation properties. Subsequently, the fermented milk was centr-ifuged at 8000×g for 5 min at 4°C, leading to the separation of curd and whey. The whey, located in the upper layer, was carefully collected and subjected to a second centrifugation at the same conditions. The whey was subsequently passed through a 0.22 µm membrane filter to prepare it for further analysis.
2.4 Acidy analysis of fermented A2 milk product
The sample’s pH was determined with a pH meter (Laqua pH1100, Horiba Scientific, Kyoto, Japan). The pH meter was calibrated with buffer solutions of pH 4.0 and 7.0. The acidity measurement using the acid-base titration method. Total of 1 mL sample diluted with 10 mL of sterile water. Afterward, 13 drops of phenolphthalein indicator were added, followed by titration of the sample with 0.05 N NaOH solution. The titration's end point was indicated by the development of a consistent pink color. Total acid was calculated according to the Eq.1.
Total acid (% lactic acid) =
[ Eq.1
2.5 Proteolytic activity assay
Proteolytic activity testing refers to the report of Celik et al. [27] with modifications. The isolate was first cultured in MRSB and incubated at 37 °C for 24 h. Cells were harvested by centrifugation (5000×g, 5 min), washed twice with phosphate-buffered saline, and adjusted to an OD600 of 0.1 ± 0.005. The resulting suspension was inoculated into 5 mL of sterile A2 milk and incubated at 37 °C for another 24 h. Following incubation, the sample was mixed with 10 mL of 0.72 N trichloroacetic acid and 1 mL of distilled water, allowed to stand for 10 min, and then filtered through Whatman No. 1 paper. From the filtrate, 5 mL was combined with 10 mL of Na2CO3–Na4P2O7 solution, after which 3 mL of diluted Folin–Ciocalteu reagent was added. The mixture was stirred until a blue color appeared, centrifuged (3000×g, 3 min), and the clear supernatant was analyzed spectrophotometrically at 650 nm. Tyrosine served as the standard for constructing the calibration curve to determine proteolytic activity.
2.6 Preparation of whey fraction by ultrafiltration
Purification using ultrafiltration refers to Yusuf et al. [26]. Whey was fractionated using a centrifugal tube containing 3 and 10 kDa filter membranes. Treatment carried out: 15 mL of supernatant was put into a 10 kDa centrifuge tube, then centrifuged (8000×g; 5 min). From this process, 2 parts of the solution will be obtained, namely those that pass the filter and those that do not pass the filter. The solution that passes the filter will contain molecules measuring ≤10 kDa and those that do not pass the filter will contain Protein with molecular weight >10 kDa. For the solution that passes the filter, it is put into a 3 kDa centrifuge tube and centrifuged again. So that 3 solutions are obtained: (1) supernatant contains molecules measuring >10 kDa, (2) supernatant contains molecules measuring ≤10 to >3kDa, and (3) supernatant contains molecules measuring ≤3 kDa. All solutions are added with distilled water up to 15 mL according to the initial volume. After that it is ready for further analysis.
2.7 The α-glucosidase inhibitor assay
The α-glucosidase inhibitor assay refers to the study of Son et al. [28]. The reaction mixture in this assay consisted of a blank control (B0), a blank (B1), a sample control (S0), and a sample solution (S1). The S1 solution was prepared by combining 150 µL of PBS (pH 7.4), 75 µL of 20 mM p-nitrophenyl α-D-glucopyranoside, and 25 µL of sample, followed by incubation at 37 °C for 10 min. Subsequently, 50 µL of 0.2 U.mL-1 α-glucosidase enzyme was added and the mixture was incubated again at 37 °C for 10 min. In contrast, the S0 solution was prepared without the addition of the α-glucosidase enzyme. To terminate the reaction, 1 mL of 0.1 M Na2CO3 was added to each mixture. The absorbance of all solutions was then recorded at 405 nm using a microplate reader (iMark, Bio-Rad Co.). After the absorbance value was obtained, the percentage of α-glucosidase inhibitor activity was calculated according to the Eq.2.
α-glucosidase inhibitor activity (%) =
Eq. 2
where B0 is blank control solution, B1 is blank solution, S0 is sample control solution, and S1 is sample solution.
2.8 The α-amylase inhibitor assay
The α-amylase inhibitory assay was performed following the method of Sato et al. [29]. Briefly, 50 µL of sample was mixed with 50 µL of α-amylase solution (1 mg.mL-1) and incubated at 25 °C for 10 min. Next, 50 µL of starch solution (20 mg.mL-1 in PBS) was added and the mixture was incubated at 37 °C for another 10 min. Subsequently, 100 µL of 3, 5-dinitrosalicylic acid was added, and the mixture was heated at 95 °C for 5 min. After cooling, 1500 µL of distilled water was added, and the mixture was centrifuged at 5000×g. An aliquot of 200 µL was transferred into a 96-well plate, and absorbance was measured at 540 nm. Acarbose at 50 ppm served as the positive control. The percentage of α-amylase inhibitory activity was then calculated using the Eq.3.
α-amylase inhibitor activity (%) =
[(Ac - (As - Ab)) ÷ Ac] × 100% Eq. 3
where As is the absorbance of the sample. Ab is the absorbance of the blank. Ac is the absorbance of the acarbose (control).
2.9 The DPP4 inhibitor assay
The DPP4 inhibitory activity was assessed using a modified version of the method described by Yan et al. [5]. In a 96-well microplate, 25 µL of gly-pro-p-nitroanilide (0.2 mM) was combined with either 25 µL of bacterial sample, PBS (as control), or sitagliptin (as reference inhibitor), followed by preincubation at 37 °C for 10 min. Subsequently, 50 µL of DPP4 enzyme (0.01 U.mL-1) was added, and the mixture was incubated at 37 °C for 60 min. The reaction was terminated by adding 100 µL of sodium acetate buffer (1 M, pH 4.0). Fluorescence intensity was then recorded using a microplate reader (Varioskan™ LUX, Thermo Fisher, Massachusetts, USA) at an excitation wavelength of 360 nm and an emission wavelength of 460 nm. All measurements were performed in triplicate, and absorbance values were corrected against blanks prepared by substituting DPP4 with Tris–HCl buffer (100 mM, pH 8.0). Negative controls (without DPP4 activity) and positive controls (DPP4 activity without inhibitor) were also included, using Tris–HCl buffer in place of the sample or the enzyme, respectively. The percentage inhibition of DPP4 activity was then calculated using the Eq. 4.
DPP4 inhibitor activity (%) =
[ Eq. 4
2.10. Identification of peptides
The method used refers to Yusuf et al. [26] with modifications. Identification using Thermo Scientific™ Dionex™ Ultimate 3000 RSLCnano UHPLC coupled with Thermo Scientific™ Q Exactive™ High Resolution Mass Spectrometer. Nano Pump: A= Water + 0.1% Formic Acid and B= Acetonitrile + 0.1% Formic acid. Analytical Column: EASY-Spray column, 15 cm × 75 μM ID, PepMap C18, 3 μm. Flow: 100 µL.min-1. Injection volume: 5 µL. Run time: 60 minutes gradient. Full MS at 70,000 FWHM Resolution. Data Dependent MS2 at 17,500 FWHM. Easy Nano Spray Ionization and positive mode. Protein identification by Thermo Scientific™ Proteome Discoverer 2.2 Software.
2.11 Molecular docking simulation and binding site
Molecular docking analysis was performed using the ClusPro webserver, which provides a dedicated peptide-protein docking feature. In this study, DPP4 was designated as the receptor protein, while the peptide VLVLDTDYK functioned as the ligand. The peptide-protein complex was selected based on the largest cluster generated by ClusPro’s docking algorithm [30]. The resulting complex structure was visualized using PyMOL and LigPlot+, and the binding affinity was evaluated using the PRODIGY webserver [31,32].
2.12 Statistical analysis
Each analysis was conducted in triplicate, and the results are presented as mean ± standard deviation. Data were statistically analyzed using SPSS software (SPSS Inc.) through one-way analysis of variance (ANOVA) followed by Duncan’s multiple range test (DMRT). A p-value of less than 0.05 (P < 0.05) was considered statistically significant.
- Results and Discussion
3.1 Profile of pH value and total acidity
The strains L. rhamnosus RAL27, Limosilactobacillus fermentum RAL29, and L. rhamnosus RAL43, when inoculated into A2 milk, exhibited normal growth. This was evidenced by their viability, which ranged from 8.68 to 9.11 log CFU.mL-1, post-fermentation pH values ranging from 4.43 to 4.66, pH changes (ΔpH) between 2.14 and 1.37, and titratable acidity between 0.65% and 0.74% (Table 1). All of these parameters are consistent with typical characteristics of fermented milk products. Comparison with previous studies [26,32,33] revealed no significant differences, with pH values after 24 hours of LAB incubation typically ranging from 3.8 to 4.8 and titratable acidity from 0.6% to 1.6%. These results confirm that A2 milk serves as an appropriate substrate for the growth of all three LAB isolates and holds potential as a starter culture for producing fermented milk or yogurt-like functional products. To assess whether the three LAB isolates could hydrolyze A2 milk proteins, their proteolytic activity was measured. The results showed that L. rhamnosus RAL27, L. fermentum RAL29, and L. rhamnosus RAL43 exhibited proteolytic activity in the range of 0.16 to 0.21 mg tyrosine.mL-1 after 24 h at 37°C (Table 1). Compared to the findings of Celik et al. [27], who reported proteolytic activity between 0.05 and 0.2 mg tyrosine.mL-1 after 36–72 hours of incubation, the values observed in this study are relatively high, especially considering the shorter fermentation time.
3.2 Proteolytic activity
This study did not include experiments to determine the optimal time and temperature for achieving maximum proteolytic activity. However, 37°C is widely recognized as the optimal temperature for LAB growth [33]. Nielsen et al. [25] also noted that the level of proteolytic activity does not necessarily correlate with the production of bioactive peptides, as specific bioactivities depend more on cleavage at the correct peptide bonds. In fact, excessively high proteolytic activity can be undesirable, as it may lead to complete breakdown into free amino acids rather than the formation of short-chain peptides. For LAB strains selected as starter cultures, a moderate level of proteolytic activity is considered desirable, since excessive protein breakdown can lead to the production of biogenic amines (organic molecules formed through amino acid decarboxylation) that may provoke allergic responses [27].
LAB can produce proteolytic enzymes both intra-cellularly and extracellularly, depending on substrate availability and favorable environmental conditions [34-36]. The nutrition label of the A2 milk used in this study indicated a protein content of 3 g.100 mL-1, which is presumed sufficient to activate the LAB proteolytic system. This system involves transport mechanisms that facilitate the uptake of nitrogen sources by the cell, proteinases that initially hydrolyze milk proteins into peptides, and peptidases that subsequently degrade these peptides into smaller fragments and free amino acids [35,37,38]. This explains the ability of LAB to efficiently degrade proteins such as casein, resulting in the release of short-chain peptides and amino acids used in bacterial metabolism.
3.3 Enzyme inhibitory activity of fermented whey fractions
After the fermentation process was completed, the whey fractions from each isolate were collected and analyzed for their enzyme inhibitory activity. The results showed that all three isolates exhibited inhibitory effects against α-glucosidase, α-amylase, and DPP4 enzymes, with varying degrees of effectiveness (Figure 1). The measurements were validated using positive controls: acarbose for α-glucosidase and α-amylase, and sitagliptin for DPP4.
RAL43 exhibited the strongest α-glucosidase inhibitory activity (66.35%), followed by RAL27 (60.18%) and RAL29 (58.07%). Although these values were significantly lower than the inhibition level of 200 ppm acarbose (89.25%), RAL43 exhibited significantly stronger inhibition than RAL27 and RAL29. For α-amylase inhibition, RAL43 again showed the highest activity at 68.87%, followed by RAL27 (65.69%) and RAL29 (64.71%), with RAL43 differing significantly from the other two isolates. In contrast, DPP4 inhibitory activity was relatively low in all samples, with RAL43 at 10.69%, RAL29 at 9.02%, and RAL27 at only 1.67%. Once again, RAL43 exhibited significantly greater inhibition compared to the other isolates.
These results indicate that RAL43 demonstrated the highest inhibitory activity across all three digestive enzymes tested. Interestingly, although RAL43 and RAL27 both belong to the species L. rhamnosus, their enzyme inhibitory performance varied considerably. This difference may be attributed to variations in their proteolytic activity. Notably, RAL43 exhibited higher proteolytic activity than RAL27, which could have led to the release of more bioactive peptides with enzyme-inhibitory properties. Yan et al. [5] also emphasized that the bioactivity of microbial strains is often strain-specific and not solely determined by species identity
The observed DPP4 inhibition (~10%) is modest compared to pharmaceutical standards. However, the purpose of this study was to provide proof-of-concept that A2 milk-derived peptides generated by L. rhamnosus RAL43 can exert measurable inhibitory activity. Importantly, the same fermented product demonstrated strong inhibition of α-glucosidase (66.35%) and α-amylase (68.87%), which are key enzymes in postprandial glucose regulation. This suggests that, while DPP4 inhibition alone may appear limited, the combined inhibitory effects on multiple enzymes can act synergistically to provide meaningful functional benefits. Further optimization of fermentation conditions or peptide concentration could enhance these outcomes.
3.4 Investigation of bioactive peptides with enzyme inhibitory activity
Since L. rhamnosus RAL43 exhibited the highest and significantly different enzyme inhibitory activity compared to the other two isolates, its whey fraction was selected for further analysis to identify the bioactive peptides responsible for the inhibition. The whey fraction obtained from RAL43 was processed through ultrafiltration membranes with molecular weight cut-offs of 3 kDa and 10 kDa.
The results showed that the >10 kDa fraction had the highest α-glucosidase inhibitory activity (36.33%), which was significantly different from the 10–3 kDa fraction (11.86%) and the <3 kDa fraction (9.92%). A comparable pattern was found in α-amylase and DPP4 inhibition, with the >10 kDa fraction showing the greatest activity compared to the other fractions (Figure 2). Ultrafiltration essentially separates molecules by size, which may lead to a dilution or loss of bioactivity. Notably, DPP4 inhibitory activity was undetectable in the <3 kDa fraction. These findings suggest that the majority of enzyme-inhibiting bioactive compounds in the RAL43 whey fraction are likely larger than 10 kDa.
Previous studies have frequently reported that enzyme-inhibitory bioactive peptides are generally found in lower molecular weight fractions [39–41]. The observation that the >10 kDa fraction of RAL43 whey exhibited the highest enzyme-inhibitory activity contrasts with previous reports, which generally associate bioactive peptides with lower molecular weight fractions (<10 kDa). Smaller peptides are typically considered more effective inhibitors due to their higher solubility and ability to access enzyme active sites. The unexpected activity in the >10 kDa fraction suggests that the active compounds in RAL43 whey may consist of larger peptides, protein-derived complexes, or even non-peptide molecules, indicating a distinct inhibitory mechanism compared to those commonly described in the literature.
3.5. Proteomic screening of the >10 kDa fraction by LC-HRMS
Proteomic screening of the >10 kDa fraction successfully revealed a variety of peptide fragments, each identifiable by its retention time and relative abundance. The complete data are presented in Figure 3. Each chromatographic peak corresponds to a distinct peptide component, and peak intensity directly correlates with peptide concentration in the sample. Protein profiling results are shown in Table 2, indicating that β-casein was the main source of peptides (referred to as the mother protein), along with contributions from β-lactoglobulin and α-lactalbumin. Based on the UniProt database, a total of 12 proteins were detected, each with an average molecular weight above 10 kDa. Hydrolysis of these proteins generated unique peptide sequences ranging from 200 to 800 Da, appearing at retention times between 1 and 21 minutes.
To assess the potential bioactivity of the identified peptides, we conducted an extensive search through various peptide functionality databases and scientific literature. This analysis identified five peptide fragments previously reported as DPP4 inhibitors. However, no confirmed α-glucosidase or α-amylase inhibitory peptides were detected in the dataset. This finding raises the possibility that the α-glucosidase and α-amylase inhibition observed in the RAL43 fraction may originate from non-peptide bioactive compounds, warranting further investigation into such molecules.
Additionally, 14 other peptide fragments were identified, each associated with diverse bioactivities, including hypocholesterolemic, splenocyte proliferation stimulation, antimicrobial, ACE-inhibitory, antioxidant, immunomodul-atory, opioid, antianxiety, and antithrombotic effects (Table 3). These peptides reflect the bioactive potential of L. rhamnosus RAL43 when fermented in A2 milk. Such findings highlight the potential of these peptides for use in the development of functional foods or novel, natural therapeutic agents.
3.5 Molecular docking analysis
From the proteomic screening results, five candidate peptides were identified as potential DPP4 inhibitors: TPEVDDEALEK, TPEVDDEALEKFDK, VLVLDTDYK, ILDKVGINYWLAHK, and VGINYWLAHK. Among these, the peptide VLVLDTDYK was selected for molecular docking due to its moderate length (nine amino acids), making it suitable for processing with ClusPro. The peptide was input together with its motif sequence (VLVLDTD[FY]K).
Ten docking poses generated under the Balanced and VdW+Electrostatic modes are shown in Figures 4. In the balanced mode results, which shown by the orange rectangle, are successfully docked and located in DPP4 (grey cartoon) at each cluster. The first to the last cluster position order starts from the top left corner to the bottom right corner, respectively, and give 66, 53, 36, 34, 24, 23, 22, 19, 17, 16 cluster sizes, respectively. Whereas, in the Vdw+electrostatic mode results, which shown by the turqoise green rectangle, are successfully docked and located in DPP4 (grey cartoon) at each cluster. The first to the last cluster position order starts from the top left corner to the bottom right corner, respectively, and give 62, 42, 41, 38, 32, 31, 37, 25, 18, 17 cluster sizes, respectively.
The conformations with the highest probability of occurrence (located in the first cluster of each mode) were selected for further analysis. These representative complex models were then visualized using PyMOL (Figure 5), where each peptide docked at different binding locations on the DPP4 protein. Detailed residue-level interactions between the peptide and protein were analyzed using LigPlot+ (Figure 6). The medium orchid-colored stick and the purple-colored finger-like shape represent DPPIV residues as a receptor protein, while the dark-blue stick represents VLVLDTDYK peptide as a ligand. The sticks always establish hydrogen and/or salt bridge (electrostatic) interaction, while the finger-like shape corresponds hydrophobic interaction of protein residues with peptide’s atoms.
The peptide-protein complex in the Balanced mode formed hydrogen bonds through interactions between Arg40 and His533 (protein) and Leu95 and Asp96 (peptide). In the VdW+Electrostatic mode, hydrogen bonds were formed between Arg471 and Ile407 (protein) and Asp96 and Lys100 (peptide). Hydrophobic interactions in the Balanced mode involved several residues, including Pro475, Asp501, Leu504, Met509, Pro510, Ser511, Phe534, and Phe559, while in the VdW+Electrostatic mode, only Leu57 and Ser473 participated in maintaining complex stability. Notably, salt bridges were observed only in the VdW+Electrostatic complex, formed by oppositely charged residues: Asp96–Arg471 and Lys100–Glu408.
Despite these interactions, the peptide VLVLDTDYK was not found to bind at DPP4’s known hydrophobic pocket or active site. Binding affinity predictions using PRODIGY (Table 4) classified both docking models within the moderate affinity category (10⁻⁹ M > Kd ≥ 10⁻⁶ M) [55]. These findings may explain the relatively low DPP4 inhibition observed in vitro, suggesting that high inhibitory activity requires peptide binding at the designated active site.
Interestingly, the VdW+electrostatic mode, which does not utilize a reference state for conformational scoring [56], produced comparable docking results to the balanced mode. Thus, predictions from both modes are considered valid approximations. Furthermore, these results suggest that alternative or unidentified binding cavities may contribute to DPP4 inhibition, albeit with weaker activity. According to UniProt data, the detected binding regions are not located in sterically hindered areas that would preclude inhibitory function, supporting their potential involvement in modulation of DPP4 activity.
This study shows that the VLVLDTDYK peptide is able to interact with the DPP4 enzyme through multiple binding modes, but not at the main active site. This accounts for the reduced inhibitory effectiveness and suggests that effective DPP4 inhibitors need to interact directly with the enzyme’s active site to achieve stronger activity. This is in line with previous findings that binding at the active site is necessary to achieve high inhibition activity [57,58], as indeed peptides that do not bind directly to the active site tend to have lower inhibition activity, even though they may interact with other areas on the enzyme.
This study has certain limitations. The peptide analysis focused primarily on VLVLDTDYK, while other identified candidates such as TPEVDDEALEKFDK were not subjected to comparable molecular docking evaluation. This narrow focus may limit the comprehensiveness of the findings, as different peptides could exhibit distinct binding affinities, interaction profiles, or inhibitory mechanisms toward DPP4, potentially overlooking stronger or more biologically relevant inhibitors. In addition, the present work was restricted to in vitro and in silico approaches without in vivo validation, leaving questions about bioavailability, stability, and physiological efficacy unanswered. Future research should therefore extend docking and interaction analyses to all candidate peptides and include in vivo studies to confirm their biological relevance and therapeutic potential.
- Conclusion
A2 milk can be a functional substrate to produce specific bioactive peptides that inhibit enzymes responsible for blood glucose spikes associated with prediabetes and T2DM. Molecular docking results corroborated the in vitro enzyme inhibition data, confirming that peptides from A2 milk fermented with L. rhamnosus RAL43 can effectively bind and inhibit DPP4 and other glucose-regulating enzymes, thereby supporting their potential as functional food components for glycemic control. Future research should include quantitative peptide yield determination, simulated gastrointestinal digestion to assess peptide stability, bioavailability studies, and in vivo or clinical trials to evaluate their physiological impact, safety, and dietary relevance.
- Acknowledgements
The authors acknowledge LPDP and BRIN Indonesia for fully supporting this research through the Funding for Research and Innovation in Advanced Indonesia (RIIM) Batch 4.
- Declaration of competing interest
The authors report no conflicts of interest.
- Authors’ Contributions
Conceptualization: Yusuf D. Data curation: Yusuf D, Handayani R. Formal analysis: Setianingrum N. Methodology: Sulistiani. Software: Dewi FR. Validation: Kasman AAMN. Investigation: Anggadhania L. Writing - original draft: Yusuf D. Writing - review & editing: Yusuf D, Handayani R, Sulistiani, Dewi FR, Anggadhania L, Setianingrum N, Kasman AAMN.
- Using Artificial Intelligent Chatbots
In the preparation of this work, the author used ChatGPT to enhance the English language and overall readability.
- Ethical Consideration
This research did not involve human participants or animal experimentation. All experiments were performed using in vitro assays and computational analyses. Therefore, ethical approval was not required.
- Bioactive Peptides
- A2 Milk
- Antidiabetic
- Molecular Docking
ارجاع به مقاله
مراجع
Chatterjee S, Khunti K, Davies MJ. Type 2 diabetes. Lancet. 2017; 389: 2239-2251. https://doi.org/10.1016/S0140-6736(17)30058-2
Antar SA, Ashour NA, Sharaky M, Khattab M, Ashour NA, Zaid RT, Roh EJ, Elkamhawy A, Al-Karmalawy AA. Diabetes mellitus: Classification, mediators, and complications; A gate to identify potential targets for the development of new effective treatments. Biomed Pharmacother. 2023; 168: 15734. https://doi.org/10.1016/j.biopha.2023.115734
Di Stefano E, Oliviero T, Udenigwe CC. Functional significance and structure–activity relationship of food-derived α-glucosidase inhibitors. Curr Opin Food Sci. 2018; 20: 7–12. https://doi.org/10.1016/j.cofs.2018.02.008
Rochín-Medina JJ, Ramírez-Serrano ES, Ramírez K. Inhibition of α-glucosidase activity by potential peptides derived from fermented spent coffee grounds. Food Chem. 2024; 454. https://doi.org/10.1016/j.foodchem.2024.139791
Yan F, Li N, Yue Y, Wang C, Zhao L, Evivie SE, Li B, Huo G. Screening for potential novel probiotics with dipeptidyl peptidase IV-Inhibiting activity for type 2 diabetes attenuation in vitro and in vivo. Front Microbiol. 2020; 10. https://doi.org/10.3389/fmicb.2019.02855
Ankolekar C, Pinto M, Greene D, Shetty K. In vitro bioassay based screening of antihyperglycemia and antihypertensive activities of Lactobacillus acidophilus fermented pear juice. Innov Food Sci Emerg Technol. 2012; 13: 221–230. https://doi.org/10.1016/j.ifset.2011.10.008
Gong L, Feng D, Wang T, Ren Y, Liu Y, Wang J. Inhibitors of α‐amylase and α‐glucosidase: Potential linkage for whole cereal foods on prevention of hyperglycemia. Food Sci Nutr. 2020; 8: 6320–6337. https://doi.org/10.1002/fsn3.1987
Singh AK, Yadav D, Sharma N, Jin JO. Dipeptidyl peptidase (Dpp) ‐iv inhibitors with antioxidant potential isolated from natural sources: A novel approach for the management of diabetes. Pharmaceuticals. 2021; 14. https://doi.org/10.3390/ph14060586
Kashtoh H, Baek K. New insights into the latest advancement in α-amylase inhibitors of plant origin with anti-diabetic effects. Plants 2023; 12: 2944. https://doi.org/10.3390/plants12162944
Patil P, Mandal S, Tomar SK, Anand S. Food protein-derived bioactive peptides in management of type 2 diabetes. Eur J Nutr. 2015; 54: 863–880. https://doi.org/10.1007/s00394-015-0974-2
Yusuf D, Nuraida L, Dewanti-Hariyadi R, Hunaefi D. In Vitro Characterization of Lactic Acid Bacteria from Indonesian Kefir Grains as Probiotics with Cholesterol-Lowering Effect. J Microbiol Biotechnol. 2020; 30: 726–732. https://doi.org/10.4014/jmb.1910.10028
Derosa G, Maffioli P. α-Glucosidase inhibitors and their use in clinical practice. Arch Med Sci. 2012; 8: 899–906. https://doi.org/10.5114/aoms.2012.31621
Riyaphan J, Pham DC, Leong MK, Weng CF. In silico approaches to identify polyphenol compounds as α-glucosidase and α-amylase inhibitors against type-ii diabetes. Biomolecules. 2021; 11. https://doi.org/10.3390/biom11121877
Agu PC, Afiukwa CA, Orji OU, Ezeh EM, Ofoke IH, Ogbu CO, Ugwuja EI, Aja PM. Molecular docking as a tool for the discovery of molecular targets of nutraceuticals in diseases management. Sci Rep. 2023; 13: 1–18. https://doi.org/10.1038/s41598-023-40160-2
Lacroix IME, Li-Chan ECY. Food-derived dipeptidyl-peptidase IV inhibitors as a potential approach for glycemic regulation - Current knowledge and future research considerations. Trends Food Sci Technol. 2016; 54: 1–16. https://doi.org/10.1016/j.tifs.2016.05.008
Nongonierma AB, Cadamuro C, Le Gouic A, Mudgil P, Maqsood S, FitzGerald RJ. Dipeptidyl peptidase IV (DPP4) inhibitory properties of a camel whey protein enriched hydrolysate preparation. Food Chem. 2019; 279: 70–79. https://doi.org/10.1016/j.foodchem.2018.11.142
González-Montoya M, Hernández-Ledesma B, Mora-Escobedo R, Martínez-Villaluenga C. Bioactive peptides from germinated soybean with anti-diabetic potential by inhibition of dipeptidyl peptidase-IV, α-amylase, and α-glucosidase enzymes. Int J Mol Sci. 2018; 19. https://doi.org/10.3390/ijms19102883
Punaro GR, Maciel FR, Rodrigues AM, Rogero MM, Bogsan CSB, Oliveira MN, Ihara SSM, Araujo SRR, Sanches TRC, Andrade LC, Higa EMS. Kefir administration reduced progression of renal injury in STZ-diabetic rats by lowering oxidative stress. Nitric Oxide - Biol Chem. 2014; 37: 53–60. https://doi.org/10.1016/j.niox.2013.12.012
Rosa DD, Dias MMS, Grześkowiak ŁM, Reis SA, Conceição LL, Peluzio MDCG. Milk kefir: Nutritional, microbiological and health benefits. Nutr Res Rev. 2017; 30: 82–96. https://doi.org/10.1017/S0954422416000275
Rubak YT, Nuraida L, Iswantini D, Prangdimurti E. Angiotensin-I-converting enzyme inhibitory peptides in milk fermented by indigenous lactic acid bacteria. Vet World. 2020; 13: 345–353. https://doi.org/10.14202/vetworld.2020.345-353
Yusuf D, Setiarto RHB, Febrisiantosa A, Firmansyah AM, Kurniawan T, Setiyawan AI, Wahyuningsih R, Ujilestari T, Rakhmatulloh S, Rizqiati H. Modification of substrate and fermentation process to increase mass and customize physical properties of L. rhamnosus and Limosilactobacillus fermentum exopolysaccharides in kefir grain. Microbiol Biotechnol Lett. 2024; 52: 152–162. https://doi.org/10.48022/mbl.2312.12013
Yusuf D, Nuraida L, Dewanti-Hariyadi R, Hunaef D. In vitro antioxidant and α-glucosidase inhibitory activities of Lactobacillus spp. Isolated from Indonesian kefir grains. Appl Food Biotechnol. 2021; 8: 39–46. https://doi.org/10.22037/afb.v8i1.30367
Celik OF, Con AH, Saygin H, Şahin N, Temiz H. Isolation and identification of lactobacilli from traditional yogurts as potential starter cultures. Lwt. 2021; 148. https://doi.org/10.1016/j.lwt.2021.111774
Son SH, Jeon HL, Yang SJ, Lee NK, Paik HD. In vitro characterization of Lactobacillus brevis KU15006, an isolate from kimchi, reveals anti-adhesion activity against foodborne pathogens and antidiabetic properties. Microb Pathog. 2017; 112: 135–141. https://doi.org/10.1016/j.micpath.2017.09.053
Sato K, Miyasaka S, Tsuji A, Tachi H. Isolation and characterization of peptides with dipeptidyl peptidase IV (DPPIV) inhibitory activity from natto using DPPIV from Aspergillus oryzae. Food Chem. 2018; 261: 51–56. https://doi.org/10.1016/j.foodchem.2018.04.029
Kozakov D, Hall DR, Xia B, Porter KA, Padhorny D, Yueh C, Beglov D, Vajda S. The ClusPro web server for protein–protein docking. Nat Protoc. 2017; 12: 255–278. https://doi.org/10.1038/nprot.2016.169
Honorato R V., Koukos PI, Jiménez-García B, Tsaregorodtsev A, Verlato M, Giachetti A, Rosato A, Bonvin AMJJ. Structural biology in the clouds: The WeNMR-EOSC ecosystem. Front Mol Biosci. 2021; 8: 1–7. https://doi.org/10.3389/fmolb.2021.729513
Xue LC, Rodrigues JP, Kastritis PL, Bonvin AM, Vangone A. PRODIGY: A web server for predicting the binding affinity of protein–protein complexes. Bioinformatics. 2016; 32: 3676–3678. https://doi.org/10.1093/bioinformatics/btw514
García-Burgos M, Moreno-Fernández J, Alférez MJM, Díaz-Castro J, López-Aliaga I. New perspectives in fermented dairy products and their health relevance. J Funct Foods. 2020; 72: 104059. https://doi.org/10.1016/j.jff.2020.104059
Dan T, Hu H, Li T, Dai A, He B, Wang Y. Screening of mixed-species starter cultures for increasing flavour during fermentation of milk. Int Dairy J. 2022; 135: 105473. https://doi.org/10.1016/j.idairyj.2022.105473
Rakhmanova A, Khan ZA, Shah K. A mini review fermentation and preservation: role of Lactic Acid Bacteria. MOJ Food Process Technol. 2018; 6: 414–417. https://doi.org/10.15406/mojfpt.2018.06.00197
Nielsen SDH, Liang N, Rathish H, Kim BJ, Lueangsakulthai J, Koh J, Qu Y, Schulz HJ, Dallas DC. Bioactive milk peptides: an updated comprehensive overview and database. Crit Rev Food Sci Nutr. 2023; 0: 1–20. https://doi.org/10.1080/10408398.2023.2240396
Atanasova J, Moncheva P, Ivanova I. Proteolytic and antimicrobial activity of lactic acid bacteria grown in goat milk. Biotechnol Biotechnol Equip. 2014; 28(4): 1073–1078. https://doi.org/10.1080/13102818.2014.971487
Kieliszek M, Pobiega K, Piwowarek K, Kot AM. Characteristics of the proteolytic enzymes produced by lactic acid bacteria. Molecules. 2021; 26. https://doi.org/10.3390/molecules26071858
Ramadhan AR, Bachruddin Z, Widodo, Erwanto Y, Hanim C. Isolation and selection of proteolytic lactic acid bacteria from colostrum of dairy cattle. IOP Conf Ser Earth Environ Sci. 2021; 788. https://doi.org/10.1088/1755-1315/788/1/012077
Liu M, Bayjanov JR, Renckens B, Nauta A, Siezen RJ. The proteolytic system of lactic acid bacteria revisited: A genomic comparison. BMC Genomics. 2010; 11: 36. https://doi.org/10.1186/1471-2164-11-36
Law J, Haandrikman A. Proteolytic enzymes of lactic acid bacteria. Int Dairy J. 1997; 7: 1–11. https://doi.org/10.1016/0958-6946(95)00073-9
Landi N, Clemente A, Pedone P V., Ragucci S, Di Maro A. An updated review of bioactive peptides from mushrooms in a well-defined molecular weight range. Toxins. 2022; 14(2). https://doi.org/10.3390/toxins14020084
Koirala P, Dahal M, Rai S, Dhakal M, Nirmal NP, Maqsood S, Al-Asmari F, Buranasompob A. Dairy milk protein–derived bioactive peptides: Avengers against metabolic syndrome. Curr Nutr Rep. 2023; 12: 308–326. https://doi.org/10.1007/s13668-023-00472-1
Sorokina L, Matic J, Rieder A, Koga S, Afseth NK, Wilson SR, Wubshet SG. Low molecular weight peptide fraction from poultry byproduct hydrolysate features dual ACE-1 and DPP4 inhibition. ACS Food Sci Technol. 2023; 3: 2219–2228. https://doi.org/10.1021/acsfoodscitech.3c00417
Silveira ST, Martínez-Maqueda D, Recio I, Hernández-Ledesma B. Dipeptidyl peptidase-IV inhibitory peptides generated by tryptic hydrolysis of a whey protein concentrate rich in β-lactoglobulin. Food Chem. 2013; 141: 1072–1077. https://doi.org/10.1016/j.foodchem.2013.03.056
Power O, Fernández A, Norris R, Riera FA, FitzGerald RJ. Selective enrichment of bioactive properties during ultrafiltration of a tryptic digest of β-lactoglobulin. J Funct Foods. 2014; 9: 38–47. https://doi.org/10.1016/j.jff.2014.04.002
Jia C li, Hussain N, Joy Ujiroghene O, Pang X yang, Zhang S wen, Lu J, Liu L, Lv J ping. Generation and characterization of dipeptidyl peptidase-IV inhibitory peptides from trypsin-hydrolyzed α-lactalbumin-rich whey proteins. Food Chem. 2020; 318: 126333. https://doi.org/10.1016/j.foodchem.2020.126333
Pihlanto-Leppälä A, Koskinen P, Phlola K, Tupasela T, Korhonen H. Angiotensin I-converting enzyme inhibitory properties of whey protein digests: Concentration and characterization of active peptides. J Dairy Res. 2000; 67: 53–64. https://doi.org/10.1017/S0022029999003982
Nagaoka S, Futamura Y, Miwa K, Awano T, Yamauchi K, Kanamaru Y, Tadashi K, Kuwata T. Identification of novel hypocholesterolemic peptides derived from bovine milkβ-lactoglobulin. Biochem Biophys Res Commun. 2001; 281: 11–7. https://doi.org/10.1006/bbrc.2001.4298
Jacquot A, Gauthier SF, Drouin R, Boutin Y. Proliferative effects of synthetic peptides from β-lactoglobulin and α-lactalbumin on murine splenocytes. Int Dairy J. 2010; 20: 514–521. https://doi.org/10.1016/j.idairyj.2010.02.013
Sedaghati M, Ezzatpanah H, Mashhadiakbar Boojar M, Tajabadi Ebrahimi M, Aminafshar M. Plasmin-digest of β-lactoglobulin with antibacterial properties. Food Agric Immunol. 2015; 26: 218–230. https://doi.org/10.1080/09540105.2014.893998
Yamada A, Sakurai T, Ochi D, Mitsuyama E, Yamauchi K, Abe F. Antihypertensive effect of the bovine casein-derived peptide Met-Lys-Pro. Food Chem. 2015; 172: 441–446. https://doi.org/10.1016/j.foodchem.2014.09.098
Udechukwu MC, Dang C, Udenigwe CC. Identification of zinc-binding peptides in ADAM17-inhibiting whey protein hydrolysates using IMAC-Zn2+ coupled with shotgun peptidomics. Food Prod Process Nutr. 2021; 3. https://doi.org/10.1186/s43014-020-00048-4
Tonolo F, Fiorese F, Moretto L, Folda A, Scalcon V, Grinzato A, Ferro S, Arrigoni G, Bindoli A, Feller E, Bellamio M, Marin O, Rigobello MP. Identification of new peptides from fermented milk showing antioxidant properties: Mechanism of action. Antioxidants. 2020; 9: 1–24. https://doi.org/10.3390/antiox9020117
Takahashi M, Moriguchi S, Suganuma H, Shiota A, Tani F, Usui H, Kurahashi K, Sasaki R, Yoshikawa M. Identification of Casoxin C, an ileum-contracting peptide derived from bovine κ-casein, as an agonist for C3a receptors. Peptides. 1997; 18: 329–336. https://doi.org/10.1016/S0196-9781(97)00044-2
Wada Y, Lönnerdal B. Bioactive peptides released by in vitro digestion of standard and hydrolyzed infant formulas. Peptides. 2015; 73: 101–105. https://doi.org/10.1016/j.peptides.2015.09.005
Liu H, Tu M, Cheng S, Xu Z, Xu X, Du M. Anticoagulant Decapeptide Interacts with Thrombin at the Active Site and Exosite-I. J Agric Food Chem. 2020; 68: 176–184. https://doi.org/10.1021/acs.jafc.9b06450
Kastritis PL, Moal IH, Hwang H, Weng Z, Bates PA, Bonvin AMJJ, Janin J. A structure-based benchmark for protein-protein binding affinity. Protein Sci. 2011; 20: 482–491. https://doi.org/10.1002/pro.580
Chuang GY, Kozakov D, Brenke R, Comeau SR, Vajda S. DARS (Decoys As the Reference State) potentials for protein-protein docking. Biophys J. 2008; 95: 4217–4227. https://doi.org/10.1529/biophysj.108.135814
Pantaleão SQ, Maltarollo VG, Araujo SC, Gertrudes JC, Honorio KM. Molecular docking studies and 2D analyses of DPP4 inhibitors as candidates in the treatment of diabetes. Mol Biosyst. 2015; 11: 3188–3193. https://doi.org/10.1039/c5mb00493d
Meduru H, Wang YT, Tsai JJP, Chen YC. Finding a potential dipeptidyl peptidase-4 (DPP4) inhibitor for type-2 diabetes treatment based on molecular docking, pharmacophore generation, and molecular dynamics simulation. Int J Mol Sci. 2016; 17: 1–12. https://doi.org/10.3390/ijms17060920
- چکیده مشاهده شده: 327 بار
- pdf (English) دانلود شده: 188 بار