Metabolomic Profiling of Industrially Associated Lactococcus Strains for Consortia Design
Applied Food Biotechnology,
Vol. 13 No. 1 (2026),
7 December 2025
,
Page 1-11 (e2)
https://doi.org/10.22037/afb.v13i1.50214
Abstract
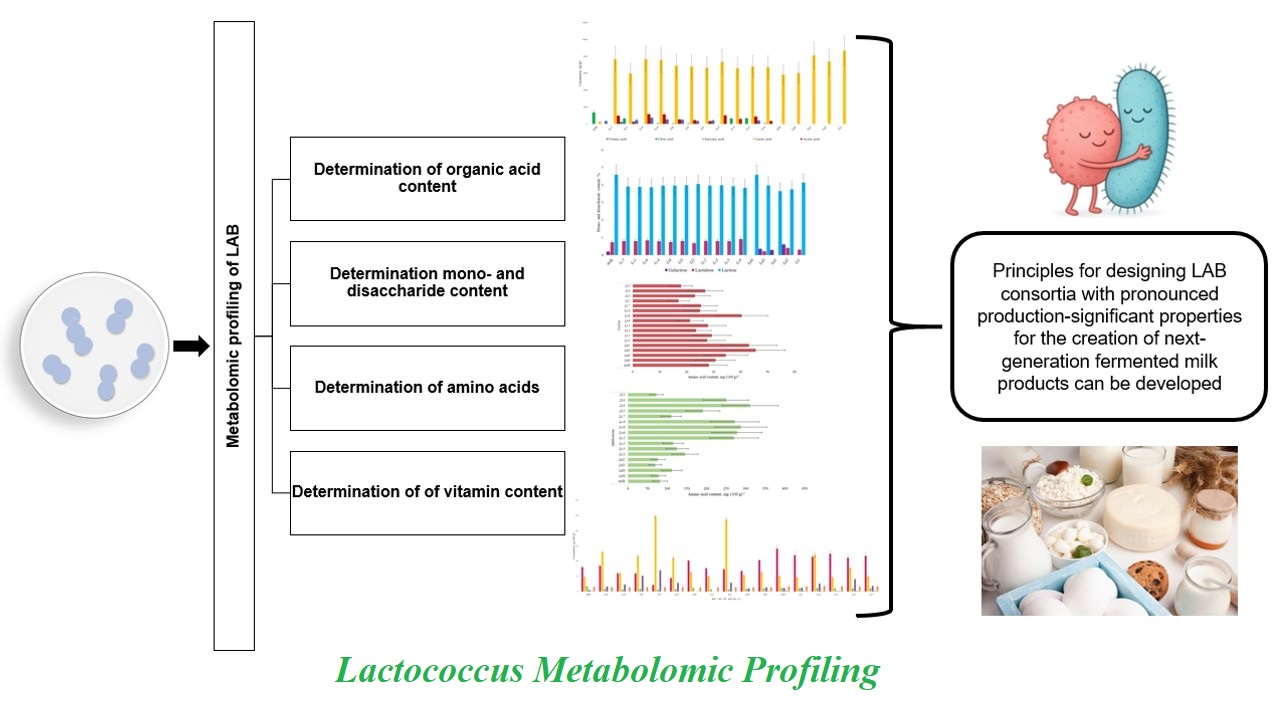
Background and Objective: The metabolomic profile represents the totality of all low-molecular-weight metabolites. It reflects the physiological state of micro-organisms and their potential for biotechnological uses. The aim of this study was to analyze the metabolomic profile of Lactococcus strains, which allowed the researchers to study the characteristics of these microorganisms. As a result, principles for designing lactic acid bacteria consortia with pronounced production-significant chara-cteristics for the creation of next-generation fermented milk products can be developed.
Material and Methods: The study microbes were Lactococcus strains. The content of organic and amino acids in the experimental samples was assessed by capillary electrophoresis. The contents of vitamins, mono- and disaccharides were assessed using high-performance liquid chromatography.
Results and Conclusion: The metabolomic profiles of 16 industrially associated Lactococcus strains were analyzed. All strains produced lactic acid, with the highest content observed in Strain Ll1, at 8677.8 mg·kg-1. The experimental samples showed a significant increase in methionine content, with the highest value of 311.2 mg·(100 g)-1 in Strain Ll3 and cystine content, with the highest value of 45.6 mg·(100 g)-1 in Strain Ld1. Strain Lc8 showed the ability to synthesize methionine and cystine, which might be promising in cheese production, since these sulfur-containing amino acids contribute to the aroma and flavor of fermented milk products. This strain produced formic, succinic and acetic acids and could catabolize citric acid and galactose. The Strains Ld1, Ld2 and Ld3 showed galactose accumulation, indicating a lack of enzymatic activity for the oxidation of this monosaccharide. Literature data on variability in vitamin synthesis levels between the strains have been verified. Thus, only Strain Ld2 synthesized vitamin B2, the content of which significantly exceeded the control and reached 103.18 ±2.06 μg·(100 g)-1. Five strains were capable of synthesizing vitamin B6 up to 247.69 ±5.45 μg·(100 g)-1. The vitamin B9 content increased, compared to the control, in eight samples, with maximum values of 69.59 ±1.46 μg·(100 g)-1 for Strains Ll3. The dataset on lactococci metabolomic profiles can serve as a basis for the further development of methodological systems, mathematical models and algorithms for selecting strains for the creation of consortia and the production of products, including those with functional characteristics. The results can serve as a basis for the design of consortia of lactic acid bacteria in biotechnological approaches to produce novel fermented milk products with specific characteristics, including functional ones, which include positive effects on human organs when consumed regularly.
Keywords: Lactic acid bacteria, Lactococcus, Functional characteristics, Metabolomic profiles, Metabolites
- Introduction
Fermented dairy products contain various metabolites produced by starter cultures that offer health benefits to the human body. These include organic acids, exopolysacchar-ides, vitamins and nitrogenous compounds such as peptides and amino acids. Metabolites are formed as a result of the vital activity of lactic acid bacteria (LAB), which ferment lactose into lactic acid, hydrolyze proteins and lipids to peptides, amino acids and free fatty acids. Some metabolites can regulate human metabolism, decrease inflammation or include antimicrobial effects [1]. Lactococcus bacteria (Streptococcaceae family) are among industrially asso-ciated bacteria in the dairy industry. They are Gram-positive, facultative anaerobic catalase-negative cocci [2]. Their primary role in fermentation processes includes efficient conversion of lactose into lactic acid, thereby increasing medium acidity, which enhances antimicrobial characteristics and extends the shelf life of the final product [3]. Lactose metabolism occurs via the pentose phosphate pathway and/or the Embden-Meyerhof-Parnas pathway (glycolysis) through the action of the enzyme β-galactosidase, which hydrolyzes lactose into glucose and galactose. Glucose enters glycolysis, forming pyruvate, which is subsequently reduced to L-lactic acid by lactate dehydrogenase. Another major route of lactose catabolism is the pentose phosphate pathway, in which lactose is phosphorylated to lactose-6-phosphate during transport and then cleaved into glucose and galactose-6-phosphate by phospho-β-galactosidase. Then, galactose-6-phosphate is converted into tagetose-6-phosphate by isomerase, which turns into glyceraldehyde-3-phosphate and dihydroxyace-tone phosphate. Glyceraldehyde-3-phosphate enters glycol-ysis and leads to pyruvate and L-lactic acid formation [4]. The accumulation of L-lactic acid leads to a decrease in the pH of the medium, which determines the intensity and pathway of lactic acid fermentation. Other key factors affecting this process are the acidification rate and the presence of oxygen and carbon dioxide in the medium. Within the Genus Lactococcus, L. lactis subsp. lactis stands out for its higher rate of lactose utilization and the ability to ferment maltose and trehalose [2].
Moreover, acid production is indirectly linked to exopolysaccharide (EPS) production. The EPS are high-molecular-weight carbohydrate polymers (either homo or heteropolysaccharides) secreted by microorganisms into the extracellular environment. Their precursors are intermediate metabolites of glycolysis and the pentose phosphate pathway glucose-6-phosphate, fructose-6-phosphate, uridine diphosphate glucose and other nucleotide sugars. Since these intermediates serve as precursors for lactic acid, a competitive relationship for carbohydrate substrates occurs [5]. The highest activity of the enzymatic complex of EPS synthesis is achieved at lower acidity levels during lactate biosynthesis (pH 5.0–5.5) [6, 7]. The production of EPS leads to the formation of a slimy layer surrounding the cell, which forms a colloidal aggregate in the form of an amorphous substance with little or no cell adhesion or a cohesive capsule [8]. Acid formation and then EPS production by Lactococcus spp. contribute to the texture of dairy fermented products, resulting in a smooth, creamy, homogeneous consistency with possibly slight ropiness.
Cow milk contains 0.4–0.5 mg·(100 g)-1 of potassium and sodium citrates. Among Lactococcus species, only L. lactis subsp. lactis var. Diacetylactis is capable of citrate metabolism. End products of citrate metabolism include diacetyl, acetoin, 2,3-butanediol, acetaldehyde, ethanol and lactic acid. The induction of the citrate permease catalytic enzyme is caused by the presence of lactic acid rather than citrate, which is synthesized from glucose and citrate under acidic pH conditions [9]. Thus, acid formation affects citrate metabolism. The most valuable metabolic products in this biosynthesis are diacetyl and carbon dioxide, as they improve the texture and flavor of fermented dairy products.
Lactococcus species include a limited ability to synthesize amino acids de novo from inorganic carbon sources and are therefore auxotrophic. However, fermented dairy products can be enriched with amino acids through the proteolysis of milk caseins [10]. Extracellular proteinases of Lactococcus hydrolyze caseins into oligopeptides, which are then transported into the cell and cleaved by intracellular peptidases into amino acids [11]. The activity of proteolytic enzymes and transport systems depends on the availability of nitrogen, acidity and the content of free amino acids and is controlled by regulatory systems. Particularly, L. lactis subsp. cremoris cleaves β-casein with the similar protease specificity as L. lactis subsp. lactis, but has less active proteolysis [12]. Amino acids are used for cellular biosynthesis and as precursors of biologically active compounds. Several amino acids are degraded to volatile compounds (aldehydes, ketones and alcohols), which can affect the sensory characteristics of fermented dairy products [12, 13]. Partial hydrolysis of milk proteins by lactococci, with lactose reduction, enhances digestibility in the human gastrointestinal tract (GIT) and may decrease the risk of intolerance, compared to raw milk.
Amino acids play a critical role in the synthesis of the bacteriocin of nisin (a polycyclic peptide composed of 34 amino acid residues, including the non-proteinogenic amino acid lanthionine) which is active against a broad spectrum of Gram-positive bacteria [14]. Studies have shown that the presence of amino acids such as alanine, isoleucine, serine, glutamic acid, tyrosine and tryptophan in the culture medium enhances bactericidal activity against strains of Escherichia coli and Staphylococcus aureus [14, 15]. Regarding vitamin biosynthesis, Lactococcus species are largely auxotrophic due to their adaptation to nutrient-rich environments such as milk, where several vitamins are readily available. Nevertheless, certain strains have the ability to synthesize B-group vitamins, such as folates (B9) and riboflavin (B2) [16]. Folate production levels vary significantly between the strains. Efforts to enhance B9 biosynthesis in LAB have involved optimized cultivation conditions and genetic modification through strain selection and metabolic engineering [17]. The ability of Lactococcus strains to produce riboflavin is limited and highly strain-dependent. Some strains function as “riboflavin producers” and are used to enrich dairy products with improved nutritional value, as well as imparting a yellowish color to whey and cheeses [18, 19].
The metabolomic profile reflects the key characteristics of the microorganism that determine its technological potential in dairy production. Industrially associated characteristics of strains include technological significance in the production of dairy products (e.g., fermentation time and viscosity) Understanding the genetic and regulatory mechanisms underlying metabolite biosynthesis pathways in Lactococcus species enables the targeted selection of starter cultures and development of microbial consortia, which is critical for creating novel biotechnological products. The aim of this study was to analyze the metabolomic profile of Lactococcus strains, which allowed the researchers to study the characteristics of these microorganisms. As a result, principles for designing LAB consortia with pronounced production-significant charact-eristics for the creation of next-generation fermented milk products can be developed.
- Materials and Methods
2.1. Experimental samples
The microorganisms of this study were Lactococcus strains from the collection of the All-Russian Research Institute of the Dairy Industry (“VNIMI”), selected based on an evaluation of the most important characteristics for dairy processing, including the viability after long-term storage, acidification rate, titratable acidity and pH, apparent viscosity and sensory characteristics. The selected strains included four strains of L. lactis subsp. lactis biovar diacetylactis (Ld1, Ld2, Ld3 and Ld4), eight strains of L. lactis subsp. cremoris (Lc1, Lc2, Lc3, Lc4, Lc5, Lc6, Lc7 and Lc8) and four strains of L. lactis subsp. lactis (Ll1, Ll2, Ll3 and Ll4). Before use, the strain was preserved at -80 °C under glycerol. The strain was then cultured in milk at 30 °C. A second-generation culture was used. The initial cell concentration was 107 CFU·ml-1. To obtain experimental samples, sterile skimmed milk (the "Standard" brand Complimilk, Belarus) was fermented with the test strain at a concentration of 5% and incubated at (30 °C ±2) until a clot formed (рН 4.5 ±0.2). The control was sterile skim milk. The milk was prepared by dissolving 90 g of dry skim milk in 1000 ml of tap water and sterilized by autoclaving using N-Bioteck sterilizer, Korea, at 121°C for 3 min. The initial pH was 6.8 and was set in a thermostat with the experimental samples at 30 °C. A limitation of this study was the number of strains (40). The study was limited to assessing the characteristics of representatives of a genus, lactococci, the most common type of LAB used in food production. The studied strains were isolated from homemade fermented products and natural sources in a temperate climate zone. Moreover, the study did not include genetically modified strains.
2.2. Assessment of organic and amino acids
The content of organic acids in the experimental samples was assessed using capillary electrophoresis and "Kapel 205" system, equipped with a spectrophotometric detector and a quartz capillary (75-µm inner diameter, 60-cm total length). Samples were pre-diluted with distilled water. The buffer electrolyte was prepared using benzoic acid, diethanolamine, cetyltrimethylammonium bromide and Trilon B. Separation was carried out at 20 kV with UV detection at 254 nm. Electropherograms were processed using "Elforan" software.
For the assessment of amino acid composition, the samples underwent acid hydrolysis and, alkaline hydrolysis for tryptophan, to convert protein-bound amino acids into free forms. All amino acids, except tryptophan, were derivatized into phenylisothiocarbamyl derivatives and quantified via capillary electrophoresis. Tryptophan was assessed directly without derivatization using borate buffer, +25 kV voltage and UV detection at 219 nm. Glutamic acid, aspartic acid and cystine were analyzed in phosphate buffer with β-cyclodextrin under +25 kV voltage, 50 mbar pressure and UV detection at 254 nm. The rest of amino acids (arginine, lysine, tyrosine, phenylalanine, histidine, leucine and isoleucine, methionine, valine, hydroxyproline, proline, threonine, serine, alanine and glycine) were assessed using similar method without pressure. All electropherograms were processed using "Elforan" software. For such amino acids as tryptophan, glutamic acid and glutamine, and aspartic acid and asparagine, the associated error was ±20%; for arginine, tyrosine, phenylalanine, histidine and methionine, the associated error was ±23%; for lysine, leucine and isoleucine, valine, proline, threonine, serine, alanine and glycine, the associated error was ±18%; for cystine and cysteine, the associated error was ±24%; and for organic acids, the associated error was ±20%.
2.3. Assessment of mono and disaccharide contents
The contents of mono and disaccharides in the experimental samples were assessed using high-perfor-mance liquid chromatography (HPLC) in accordance with GOST 54760-2011, “Component milk products and infant milk products. Assessment of mono and di-sugars mass concentration using HPLC method.” The analysis was carried out using MAESTRO liquid chromatograph (INTER-LAB, Russia), with a Zorbax carbohydrate analysis column (5 μm, 4.6 × 250 mm; Agilent Technologies, USA), CAU-X-320 electronic analytical balance (CAS, Korea), variable-volume single-channel pipette (100–1000 μl; BIOHIT, Finland; Sartorius Biohit Liquid Handling, Germany) and Sigma 1-14 microcentrifuge (Sigma Laborz-entrifugen, Germany). The associated measurement error was ±12%.
2.4 Assessment of vitamins
Vitamin C was assessed using Agilent 1260 chromato-graphy system equipped with a gradient 4-channel pump, a diode array detector, a column thermostat and an automatic sample delivery system. The associated measurement error was ±34%. The analyte was extracted by diluting the sample aliquot 1:2 with an extracting solution (3% metaphosphoric acid). It was vortexed and centrifuged at 10.850× g. The supernatant was filtered through a 0.22-μm filter. The resulting filtrate was used for analysis. Separation of components was carried out by reversed-phase chromate-graphy on an Agilent Extend-C18 4.6*250 mm column. Vitamin C was eluted under isocratic conditions using mobile phase of 30 mM phosphate buffer, pH 2.6, at a flow rate of 1 ml·min-1. Detection was carried out using diode array detector at 254 nm.
B-group vitamins were assessed using HPLC with an Agilent 1260 Infinity II HPLC device combined with an Ultivo Triple Quad LC/MS mod. 6465, Singapore, Agilent Technologies and an Agilent 1260 Infinity II HPLC device with a diode array detector G7115A. Chromatographic separation was carried out using Agilent InfinityLab 120 Poroshell 120 Phenyl-Hexyl column (3.0 × 100 mm, 2.7 μm). For sample preparation, 1 g of the sample was mixed with 4 cm³ of deionized water, vortexed and then 5 cm³ of acetonitrile and 0.1 g of ascorbic acid were added to the mixture. The mixture was re-vortexed, ultrasonicated for 30 min and centrifuged at 3.650× g for 10 min. The sample was stored at -4 to -6 °C and filtered through a 0.22-μm filter.
2.5. Statistics
The reliability of data was verified through the execution of experiments in at least three independent replicates. Furthermore, MS Office Excel 2016 was used for data analysis and graph creation. Experimental data were presented in“mean value ± measurement error (relative)”. The data on vitamins were present as mean ± standard deviation. To calculate the significance of differences, ANOVA statistical method was used, with the accepted limitation of statistical significance of differences between options at p ≤ 0,05. The study was carried out using equipment of the Collaborative Center of the All-Russian Dairy Research Institute (CKP “VNIMI”).
- Results and Discussion
3.1. Assessment of amino acids
Amino acids are promising metabolites for biotech-nological uses. These compounds are used as artificial sweeteners, flavoring agents and feed additives and for pharmaceutical purposes [19, 20]. During the production of fermented dairy products, specific amino acids generated via casein proteolysis are responsible for the formation of thiols, alcohols, esters and aldehydes, contributing to a broad range of flavors [20]. Sulfur-containing amino acids play a critical role in the development of sensory charac-teristics, as well as in the antioxidant activity of fermented dairy products [21]. In the experimental samples containing the industrially associated Lactococcus strains, the amino acids were assessed. Compared with the control (milk), a significant increase in the content of methionine and cystine was detected in the experimental samples. The results are presented in Figure 1.
A significant increase in amino acid content relative to the control was observed for methionine (Figure 1A) in Strains Lc2, Lc6, Lc8, Lc4, Ll2, Ll3, Ll4, with the highest content in Strain Ll3 at 311.2 mg·(100 g)-1. The sulfur-containing amino acid methionine plays a role in the initiation of transcription and is critical in various methyltransferase reactions. Aromatic compounds such as 3-methylbutanal, methanethiol, dimethyl sulfide, 2-methylpropanol and dimethyl trisulfide are synthesized from methionine [22]. Among these, methanethiol is particularly important for its contribution to the desirable flavor of cheddar cheese, especially in combination with diacetyl and butyric acid [23]. Increase in cystine content (Figure 1B) was observed in Strains Ld1, Ld2 and Lc8, with the highest content in Strain Ld1 [45.6 mg·(100 g)-1], compared to the control. Cystine is the oxidized form of cysteine, which serves as a key metabolite for synthesizing most sulfur-containing cellular compounds. Cysteine is involved in protein folding, assembly and stability through the formation of disulfide bonds. Additionally, cysteine-containing proteins such as thioredoxin and glutathione play essential roles in protecting cells against oxidative stress [24]. The strain with the conditional designation (Lc8) is capable of synthesizing methionine and cystine. The levels of other amino acids were similar to or less than those of the control, suggesting their active utilization by microorganisms during growth.
3.2. Assessment of organic acid content
One of the fundamental criteria for selecting Lactococcus strains for starter culture consortia is their ability to produce acid. Organic acids such as acetic, lactic, citric, succinic and propionic acids are produced by LAB during carbohydrate metabolism. As intermediates in metabolic pathways, these acids enhance the taste of products and extend shelf life through their antimicrobial and antioxidant characteristics; thereby, increasing consumer value [25,26]. The results of assessment of the content of organic acids are presented in Figure 2.
All the samples showed a significant increase in lactic acid content, compared to the control (Figure 2). The highest levels were observed in Strains Ld1 and Ll1, reaching 8116.5 and 8677.8 mg·kg-1, respectively. Lactic acid synthesis indicates the β-galactosidase enzymatic activity of these microorganisms. Lactic acid in fermented milk products acts as an acidity regulator and preservative. When ingested, lactic acid accelerates metabolism, decreases the number of pathogenic bacteria and also has antioxidant characteristics.
Only trace quantities of citric acid were detected in Samples Lc1, Lc3 and Lc5, which indicated the presence of a citrate pathway for the breakdown of this substrate. The consumption of citric acid depends on the presence of genes encoding the enzymes citrate permease and citrate lyase, which ensure the transport of citrate into the cell and its catabolism into oxaloacetate [27]. As a result of breakdown by lactococci, gas formation occurs in dairy products. In the production of certain products such as cheeses, this is addressed as a positive phenomenon, leading to the formation of "eyes." However, if its presence is assessed, this can lead to product defects. The breakdown of citric acid by these microorganisms leads to the formation of acetate, lactic acid, diacetyl and acetoin. Diacetyl and acetoin create flavor and aroma in food products, which is promising for the production of starters for butter, sour cream, yogurt and certain types of cheese.
Significantly, formic acid was detected in Strains Lc1, Lc2, Lc4, Lc6, Lc7, Lc8, Ll2, Ll3 and Ll4. In Lactococcus, formic acid is produced primarily by the pyruvate-formate lyase enzyme, which catalyzes the breakdown of excess pyruvate into acetyl-CoA and formate, which is the initial step in the formation of mixed-acid products. In the L. lactis genome, the genes responsible for this pathway are pfl, encoding pyruvate-formate lyase and pflA, a formate activator. Formate is formed with acetyl-CoA, which is consumed by the cell for energy needs. Formate itself can be excreted from the cell to maintain pH and redox balance [28]. In fermented dairy products, formic acid has little effect on the taste of the product, but it can serve as a preservative to preserve nutrients and it also has a bactericidal effect against pathogens.
Succinic acid was detected in Strains Lc4, Lc8, Ll3 and Ll4, with contents of 83.5, 89.9, 74.9 and 63.3 mg·kg-1, respectively. This acid is synthesized via heterofermentative lactic acid fermentation alongside lactic acid. Succinic acid is formed upon activation of the reductive branches of the tricarboxylic acid cycle. In Lactococcus strains, the complete tricarboxylic acid cycle is inactive, but succinate can be synthesized through carboxylation of C3–C4 units as pyruvate or phosphoenolpyruvate is carboxylated to oxalo-acetate by pyruvate carboxylase or phosphoenolpyruvate carboxy-lase and then oxaloacetate is sequentially reduced to malate and ultimately converted to succinate [29]. In the human body, succinic acid promotes the synthesis of propionic and butyric acids in microbiota.
Acetic acid production was observed in many strains, with the highest contents recorded in Strain Lc4 and Lc6 at 1148.7 and 1129.8 mg·kg-1, respectively. Acetate is formed during mixed fermentation from acetyl-CoA by phospho-acetyltransferase and acetate kinase. This results in the generation of energy in the cell in the form of ATP [30]. Acetic acid enhances the sensory characteristics of the fermented dairy products and contributes to microbial safety and shelf-life extension due to its antimicrobial activity.
3.3. Assessment of mono and disaccharide contents
Lactose, the primary carbohydrate in milk, is a disaccharide composed of glucose and galactose residues, hydrolyzed by lactase and β-galactosidase. Lactose metabolism in Lactococcus differs from that of other LAB by allowing simultaneous catabolism of glucose and galactose [9]. Galactose participates in the formation of glycoproteins and glycolipids, which are essential for cell-cell communication. However, its presence in food poses risks for individuals with galactosemia.
Another important disaccharide is lactulose, formed from lactose during high-temperature treatment of milk through isomerization involving hydrogen transfer from the second to the first carbon atom in the glucose residue. Lactulose content in milk serves as an indicator of thermal processing [31]. It is composed of D-fructose and D-galactose linked via a β-1,4-glycosidic bond and shows prebiotic effects at low concentrations. Previous research [32] demonstrated that the presence of lactulose improves the viability of starter cultures during storage, freezing and freeze-drying.
The analysis of monosaccharides and disaccharides (Figure 3) revealed a slight decrease in lactose content, compared to milk in all experimental samples. Particularly, lactulose was detected in all samples except Strains Ld1 and Ld4, including control. A decrease in lactulose content was observed in Samples Ld2, Ld3 and Ll1, compared to the control. The presence of lactulose can enhance consistency, increase starter culture viability and extend the shelf life of fermented dairy products. Galactose was detected in Strains Ld1, Ld2 and Ld3, with contents of 0.295%, 0.605% and 0.354% by mass, respectively. This indicated that the other strains—Lc1 through Lc8, and Ll1 through Ll4 and Ld4—were capable of galactose utilization. After transport into the microorganism, lactose contained in milk is hydrolyzed by β-galactosidase to glucose and galactose. Galactose can either be released by cells into the environment or undergo further metabolism into glucose-1-phosphate by four enzymes of galactokinase (GalK), hexose-1-phosphate uridine transferase (GalT), UDP-glucose-4-epimerase (GalE) and aldose-1-epimerase (GalM). These enzymes make up the Leloir pathway, whose genes can be present either alone (gal genes) or in combination with lactose metabolism genes (lac genes), making the final fermented products safer for individuals with galactosemia.
3.4. Assessment of vitamin contents
Based on the results of amino acid analysis, nine strains that showed a significant increase in methionine and cystine contents in samples were selected, compared to the control (Lc2, Lc4, Lc6, Lc8, Ll2, Ll3, Ll4, Ld1 and Ld2). These strains are of interest for possible inclusion in consortia for the development of products with specific functional characteristics. The selected samples were additionally assessed for vitamin C and vitamin B group (B2, B6, B7 and B9) contents. The results of vitamin C assessment in the samples are presented in Table 1.
As the studies have shown, the content of vitamin C in the samples did not exceed the control levels. The results of the assessment of B vitamins in the samples are presented in Table 2.
A number of foods, including milk, contain vitamin B2 (riboflavin). It is a key enzyme in redox reactions and essential for hemoglobin synthesis, maintaining the normal condition of the GIT mucosa and supporting nervous system function [33]. The ability of lactococci to synthesize riboflavin is limited, but some strains produce the compound in significant quantities [18]. In this study, only Strain Ld2 showed an increased level of vitamin B2 [103.18 μg·(100 g)-1], compared to the control [79.97 μg·(100 g)-1]. In the other samples, the content of riboflavin was at the control level or decreased.
Vitamin B6 plays a critical role in the normal functioning of the human body systems, including the digestive, cardiovascular and immune systems. It participates in hundreds of biochemical reactions, affecting metabolism. It is involved in the synthesis of neurotransmitters such as serotonin and dopamine, improves cognitive functions and promotes a positive mood [34]. It affects collagen synthesis and regulates hormonal balance in women. In the samples of industrially associated Lactococcus strains Lc6, Ll4, Ll3, Ll8 and Ll2, a significant content of vitamin B6 was detected, exceeding the control by 3–5 times. The vitamin B7 content in all samples was lower than that in the control.
Analysis of vitamin B9 content showed a significant increase in eight out of nine samples, ranging from 10.02 to 69.59 μg·(100 g)-1, compared to 4.37 μg·(100 g)-1 in the control group. Significantly, the Lc2 strain contained less than 1 μg·(100 g)-1 of vitamin B9. Folic acid (vitamin B9) is essential for appropriate cell growth and division, acting as a coenzyme in nucleic acid synthesis. Folic acid deficiency can lead to serious fetal development disorders in pregnant women [35]. Vitamin B9 plays a critical role in the conversion of homocysteine, a substance associated with an increased risk of cardiovascular disease, into methionine. This compound is necessary for the synthesis of several neurotransmitters, prevents neurological damage, improves cognitive function and decreases the risk of certain types of cancer (breast, cervical and intestinal cancers) [36]. Receiving vitamins through foods is more physiologically beneficial for the human body than introducing chemically synthesized preparations. A comparative assessment of lactococci from the VNIMI collection, stored since 1960, was carried out for the regenerative capacity of LAB strains, as well as for the production-significant characteristics of these microorganisms such as fermentation activity, active and titratable acidity, viscosity and organoleptic indicators. Of the 40 Lactococcus strains, 39 showed metabolic activity regardless of storage time [37].
- Conclusion
The study included metabolomic profiling of industrially associated Lactococcus strains based on the analysis of amino acids, organic acids, monosaccharides, disaccharides and vitamins. All strains produced lactic acid, with the highest content observed in Strains Ld1 and Ll1, at 8116.5 and 8677.8 mg·kg-1, respectively. The experimental samples showed a significant increase in the content of methionine, with the highest content in Strain Ll3 and cystine in Strain Ld1. The content of other amino acids was similar to the control level or decreased. Strain Lc8 showed the ability to synthesize methionine and cystine, which may be promising in cheese production, since these sulfur-containing amino acids contribute to the aroma and flavor of fermented milk products. This strain produced formic, succinic and acetic acids and could catabolize citric acid and galactose. The Strains Ld1, Ld2 and Ld3 showed galactose accumulation, indicating lack of enzymatic activity for the oxidation of this monosaccharide. Literature data on significant variability in vitamin synthesis levels between the strains have been verified [18,20]. Thus, only Strain Ld2 synthesized vitamin B2, the quantity of which significantly exceeded that in the control and reached 103.18 ±2.06 μg·(100 g)-1. Five strains were capable of synthesizing vitamin B6 up to 247.69 ±5.45 μg·(100 g)-1. The vitamin B9 content increased in eight of the nine samples, with maximum values of 69.59 ±1.46 and 51.16 ±1.28 μg·(100 g)-1 for the strains Ll3 and Ll4, compared to the control. For further formation of consortia, strains Lc1, Lc8, Ll2, Ll3, Ld1 and Ld2 were selected. The results can serve as a basis for the design of consortia of LAB in biotechnological approaches to produce novel fermented milk products with specific characteristics, including functional ones.
- Acknowledgements
The authors would like to thank Elena Yurova and Nikolai Zhizhin for their assistance.
- Declaration of competing interest
The authors report no conflict of interest (If authors have any kind of interest mention please clearly).
- Authors’ Contributions
Conceptualization, I.R., N.P., V.L., V.S., S.K., V.M.; methodology S.K.; validation, S.K., V.S.; formal analysis, I.R., V.M., S.K..; investigation, V.S., S.K.; data curation, S.K., V.S; writing—original draft preparation, V.S.; writing—review and editing, V.L., I.R., V.S., S.K.; visualization, V.S.; supervision, I.R. N.P.; project administration, I.R, N.P.
- Using Artificial Intelligent Chatbots
No using Artificial Intelligent Chatbots
- Ethical Consideration
The ethics committee's opinion is not required in this work
- Lactic acid bacteria
- Lactococcus
- Functional characteristics
- Metabolomic profiles
- Metabolites
How to Cite
References
1. Menard S. Lactic acid bacteria secrete metabolites retaining anti-inflammatory properties after intestinal transport. Gut. 2004; 53(6): 821-828. https://doi.org/10.1136/gut.2003.026252
2. Stoyanova LG. Isolation and identification of lactic acid bacteria lactococcus lactis subsp. Lactis with antimicrobial properties. Izvestiâ Timirâzevskoj selskohozâjstvennoj akademii. 2017; (5): 41-61. https://doi.org/10.26897/0021-342x-2017-5-41-61
3. Bagunova AV. Biologically active metabolites of lactic acid bacteria. Food processing industry. 2022; (6):21-25. https://doi.org/10.52653/PPI.2022.6.6.005
4. Kelleher P. Comparative and functional genomic analysis of dairy lactococci: introduction [PhD thesis]. Cork, Ireland: University College Cork; 2017. p. 12-92.
5. Ramos A, Boels IC, Willem, Santos H. Relationship between Glycolysis and Exopolysaccharide Biosynthesis in Lactococcus lactis. Appl Environ Microbiol. 2001; 67(1): 33-41. https://doi.org/10.1128/aem.67.1.33-41.2001
6. Looijesteijn PJ, Hugenholtz J. Uncoupling of growth and exopolysaccharide production by Lactococcus lactis subsp. cremoris NIZO B40 and optimization of its synthesis. J Biosci Bioeng. 1999; 88(2):178-82. https://doi.org/10.1016/s1389-1723(99)80198-4
7. Ayala-Hernández I, Hassan A, Goff HD, Mira de Orduña R, Corredig M. Production, isolation and characterization of exopolysaccharides produced by Lactococcus lactis subsp. cremoris JFR1 and their interaction with milk proteins: Effect of pH and media composition. Int Dairy J. 2008; 18(12): 1109-1118. https://doi.org/10.1016/j.idairyj.2008.06.008
8. Boels IC, Ramos A, Kleerebezem M, de Vos WM. Functional Analysis of the Lactococcus lactis galU and galE Genes and Their Impact on Sugar Nucleotide and Exopolysaccharide Biosynthesis. Appl Environ Microbiol. 2001; 67(7): 3033-3040. https://doi.org/10.1128/aem.67.7.3033-3040.2001
9. Kim W. The genus Lactococcus. In: Holzapfel WH, Wood BJB, editors. Lactic Acid Bacteria: Biodiversity and Taxonomy. Oxford, UK: Wiley-Blackwell; 2014. p. 429-443. https://doi.org/10.1002/9781118655252.ch26
10. Kondrotiene K, Zavistanaviciute P, Aksomaitiene J, Novoslavskij A, Malakauskas M. Lactococcus lactis in dairy fermentation—health-promoting and probiotic properties. Fermentation [Internet]. 2024; 10(1):16. Available from: https://www.mdpi.com/2311-5637/10/1/16 https://doi.org/10.3390/fermentation10010016
11. Kieliszek M, Pobiega K, Piwowarek K, Kot AM. Characteristics of the proteolytic enzymes produced by lactic acid bacteria. Molecules [Internet]. 2021; 26(7):1858. Available from: https://www.mdpi.com/1420-3049/26/7/1858 https://doi.org/10.3390/molecules26071858
12. Helinck S, Le Bars D, Moreau D, Yvon M. Ability of thermophilic lactic acid bacteria to produce aroma compounds from amino acids. Appl Environ Microbiol. 2004 Jul; 70(7):3855-3861. https://doi.org/10.1128/aem.70.7.3855-3861.2004
13. Khavkin AI, Kovtun TA, Makarkin DV, Fedotova OB. Fermented milk products and child health. Rossiyeskiy Vestnik Perinatologii i Pediatrii. 2021; 65(6):155-165. https://doi.org/10.21508/1027-4065-2020-65-6-155-165
14. Tavares TD, Ana R.M. Ribeiro, Silva C, Antunes JC, Felgueiras HP. Combinatory effect of nisin antimicrobial peptide with bioactive molecules: A review. J Drug Deliv Sci Technol. 2024; 91: 105246. https://doi.org/10.1016/j.jddst.2023.105246
15. Sybesma W. Multivitamin production in Lactococcus lactis using metabolic engineering. Metab Eng. 2004; 6(2):109-115. https://doi.org/10.1016/j.ymben.2003.11.002
16. Sybesma W, Starrenburg M, Kleerebezem M, Mierau I, Vos WM de, Hugenholtz J. Increased production of folate by metabolic engineering of Lactococcus lactis. Appl Environ Microbiol. 2003; 69(6):3069-3076. https://doi.org/10.1128/AEM.69.6.3069-3076.2003
17. LeBlanc JG, Burgess C, Sesma F, de Giori GS, van Sinderen D. Ingestion of milk fermented by genetically modified Lactococcus lactis improves the riboflavin status of deficient rats. J Dairy Sci. 2005; 88(10): 3435-3442. https://doi.org/10.3168/jds.s0022-0302(05)73027-7
18. Hirasawa T, Shimizu H. Recent advances in amino acid production by microbial cells. Curr Opin Biotechnol. 2016; 42: 133-146. https://doi.org/10.1016/j.copbio.2016.04.017
19. Pinu FR, Granucci N, Daniell J, Han TL, Carneiro S, Rocha I, et al. Metabolite secretion in microorganisms: The theory of metabolic overflow put to the test. Metabolomics. 2018; 14(4). https://doi.org/10.1007/s11306-018-1339-7
20. van Kranenburg R, Kleerebezem M, van Hylckama Vlieg J, Ursing BM, Boekhorst J, Smit BA, et al. Flavour formation from amino acids by lactic acid bacteria: Predictions from genome sequence analysis. Int Dairy J. 2002;12(2-3):111-121 https://doi.org/10.1016/S0958-6946(01)00132-7
21. Donskaya GA. Antioxidant properties of milk and dairy products: An overview. Food industry. 2020;12: 86-91. https://doi.org/10.24411/0235-2486-2020-10150
22. Amárita F. Conversion of methionine to methional by Lactococcus lactis. FEMS Microbiol Lett. 2001; 204(1):189-195. https://doi.org/10.1016/s0378-1097(01)00402-5
23. Babkina, NG. Main trends and technological aspects of semi-hard cheese production. Functions of starter and protective microflora. Cheese Butter Making. 2021;(1): 22-24.
24. Sperandio B, Polard P, Ehrlich DS, Renault P, GuédonE. Sulfur amino acid metabolism and its control in Lactococcus lactis IL1403. J Bacteriol. 2005; 187(11): 3762-3778. https://doi.org/10.1128/jb.187.11.3762-3778.2005
25. Punia Bangar S, Suri S, Trif M, Ozogul F. Organic acids production from lactic acid bacteria: A preservation approach. Food Biosci. 2022; 46: 101615. https://doi.org/10.1016/j.fbio.2022.101615
26. Coban HB. Organic acids as antimicrobial food agents: Applications and microbial productions. Bioprocess Biosyst Eng. 2020; 43(4): 569-591. https://doi.org/10.1007/s00449-019-02256-w
27. Laroute V, Tormo H, Couderc C, Mercier-Bonin M, Le Bourgeois P, Cocaign-Bousquet M, et al. From genome to phenotype: An integrative approach to evaluate the biodiversity of Lactococcus lactis. Microorganisms. 2017; 5(2). https://doi.org/10.3390/microorganisms5020027
28. Melchiorsen, RC, Jokumsen, VK, Villadsen, J, Israelsen, H, Arnau, J. The level of pyruvate-formate lyase controls the shift from homolactic to mixed-acid product formation in Lactococcus lactis. Appl Microbiol Biotechnol. 2002; 58(3): 338-344. https://doi.org/10.1007/s00253-001-0892-5
29. Lin H, San KY, Bennett GN. Effect of Sorghum vulgare phosphoenolpyruvate carboxylase and Lactococcus lactis pyruvate carboxylase coexpression on succinate production in mutant strains of Escherichia coli. Appl Microbiol Biotechnol. 2004; 67(4):515-523. https://doi.org/10.1007/s00253-004-1789-x
30. Puri P, Goel A, Bochynska A, Poolman B. Regulation of acetate kinase isozymes and its importance for mixed-acid fermentation in Lactococcus lactis. J Bacteriol. 2014; 196(7):1386-1393. https://doi.org/ 10.1128/JB.01277-13
31. Agarkova EU, Kruchinin AG, Ryazantseva KA. Modern technological approaches to fortification of dairy products. In: Innovative technologies for fortification of dairy products (theory and practice). 2016: 110-142.
32. Shpak M, Ryabtseva S, Bratsikhin A. Lactulose effect on viability of starter cultures. J Hyg Eng Des. 2019; 1, 27: 162-167.
33. Dolmatova IA, Zaitseva TN, Ryabova VF, Gorelik OV. Biological role of vitamins. Actual problems of modern science, technology and education. 2020; 11(1): 116-119.
34. Akabaeva VK. Alliance of magnesium and vitamin B6: role in clinical cardiology. In: Actual problems of medicine. 2015; 1: 19-22.
35. Eliseeva T, Mironenko A. Vitamin B9 description, benefits, effects on the body and the best sources. J Healthy Nutr Diet. 2019; 4(10): 88-100.
36. Gromova, O. A., Torshin, I. Y., Kalacheva, A. G., Semenov, V. A., & Rudakov, K. V. MexiB 6 as a result of fortification of ethylmethylhydroxypyridine succinate with magnesium and pyridoxine: Proteome effects. Neurology, Neuropsychiatry, Psychosomatics. 2016; 8(4): 38-44. https://doi.org/ 10.14412/2074-2711-2016-4-38-44
37. Semenova V, Mitrova V, Kishilova S, Rozhkova I, Petrov A. Comparative evaluation of the properties of industrially valuable strains Lactococcus. Food Process Ind. 2025; 8: 82-87. https://doi.org/10.52653/PPI.2025.8.8.022
- Abstract Viewed: 84 times
- pdf Downloaded: 55 times