A Biosafe Bacillus subtilis 168 Engineered for Hyaluronic Acid Biosynthesis
Applied Food Biotechnology,
Vol. 13 No. 1 (2026),
7 December 2025
,
Page 1-13 (e13)
https://doi.org/10.22037/afb.v13i1.51984
Abstract
Background and Objective: Hyaluronic acid is a high‑value biopolymer widely used in the food, cosmetic, therapeutic and pharmaceutical industries. This study aimed to establish a biosafe “Generally Recognized as Safe” platform for hyaluronic acid biosynthesis via engineering Bacillus subtilis 168.
Material and Methods: The hyaluronan synthase gene from Streptococcus dysgalactiae and tuaD homolog NX02_04625 from Sphingomonas sanxanigenens were selected based on their reported catalytic performance and codon‑optimized for Bacillus subtilis. The complete pHT01 vector construct harboring the two genes was commercially synthesized and sequence‑verified. A single‑gene plasmid was generated and the two plasmids were introduced into Bacillus subtilis 168 via electroporation. Hyaluronic acid production was quantified using cetyltrimethylammonium bromide assay, structurally validated by Fourier transform infrared spectroscopy and characterized by size‑exclusion chromatography.
Results and Conclusion: The dual-gene strain produced 1.20 ± 0.03 gl-¹ hyaluronic acid, achieving a 2.4‑fold increase relative to the single‑gene strain and showed a trimodal molecular‑weight profile dominated by high‑molecular-weight polymers (> 1.18 MDa, 43.45%), accompanied by medium‑range species (~0.99 MDa, 24.69%) and low‑molecular-weight-oligomers (~55 kDa, 31.86%). Collectively, these findings highlight the efficiency and biosafety of the engineered production platform and its potential for food‑grade uses, supported by the predominance of high‑molecular-weight hyaluronic acid with favorable gel‑like characteristics.
Keywords: Bacillus subtilis 168, Hyaluronan synthase gene, Hyaluronic acid production, Metabolic engineering, Strain development, UDP‑glucose dehydrogenase
- Introduction
Hyaluronic acid (HA) is a high-value, linear glycosaminoglycan renowned for its exceptional biocompatibility, viscoelasticity and hygroscopic characteristics [1]. As established in recent literature [2], HA is structurally composed of repeating disaccharide units of D-glucuronic acid (GlcUA) and N-acetyl-D-glucosamine (GlcNAc). The HA is a ubiquitous component of the extracellular matrix in vertebrates; where, it regulates cell proliferation, tissue hydration, inflammation and wound repair [3]. Due to its shear-thinning behavior and high moisture retention, HA has become essential in biomedicine (e.g. dermal fillers and drug delivery), cosmetics (e.g. anti-aging formulations) and regenerative medicine [4, 5].
The unique physicochemical characteristics of HA, including its molecular weight-dependent biological functions, support its clinical versatility [1]. High-molecular-weight (HMW) HA is generally associated with tissue homeostasis and restraint of cell proliferation/migration, whereas HA fragmentation into low-molecular-weight (LMW) species is linked to inflammatory and promigratory signaling [6]. The LMW-HA can enhance endothelial angiogenic responses and migration in vitro [7], supporting the use of tailored HA preparations for various therapeutic uses.
In comparative studies using mouse models of persistent inflammation, immunosuppression and catabolism syndrome, low (3 kDa), medium (100 kDa) and HMW (1600 kDa) HA were assessed at 30 mg kg⁻¹, revealing distinct biological responses as 1600 kDa HA significantly promoted recovery of intestinal structure and was associated with improved immune recovery while 100 kDa HA induced stronger inflammatory cell infiltration after prolonged treatment and 3 kDa HA showed comparatively milder effects [8].
The HA is a promising safe additive for functional foods. In yogurt, HA at 180, 350 and 1280 kDa showed that 350 kDa improved pH, texture and sensory quality most effectively; 1280 kDa increased stability but caused whey syneresis and 180 kDa decreased stability [9]. For sweetness at pH 4.0, HA at 100, 400 and 1090 kDa extended sweet taste duration, with 100 kDa showing the strongest longest-lasting effect via enhanced sucralose transport. Advances in sustainable production support broader uses, with HMW-HA critical for gel-like texture, stability and viscosity [10]. In meats, a 0.9% HA coating has preserved the freshness of crucian carp for 24 d [11]. The HA has formed stable complexes with fish gelatin, improving its emulsifying characteristics [12]. Products such as Cenovis HA beverage [13] and rooster comb extract in dairy products [14] demonstrate the increasing use of food-grade HA. The HA strength, biodegradability and biocompatibility include excellent potential for edible packaging. Its versatility in forming gels and complexes with neutral taste and color makes it a promising material for food coatings and edible packaging materials [15–17].
Conventional HA production methods face critical limitations. Fermentation using pathogenic Streptococcus strains (S. zooepidemicus) increases biosafety concerns and complicates the downstream processing [18, 19]. The relatively low yield of natural HA producers makes conventional production economically and industrially less feasible. This limitation has accelerated efforts to develop genetically engineered generally recognized as safe (GRAS) microbial platforms for sustainable HA synthesis [20, 21]. In recent years, Bacillus subtilis (B. subtilis) has attracted significant attentions as a microbial cell factory for the biosynthesis of valuable biomolecules and biopolymers [22-25]. Well-characterized genome, efficient protein secretion system and strong metabolic capacity has made B. subtilis a particularly attractive chassis for metabolic engineering and synthetic biology uses [22, 26].
Several studies have demonstrated the feasibility of engineering B. subtilis to produce heterologous biomolecules, highlighting its potential as a safe and scalable platform for industrial biotechnology [23, 27]. Regarding its non-pathogenic nature, endotoxin-free status and established use in industrial biotechnology, this bacterium has been engineered to produce enzymes, polysaccharides and vitamins [23, 24, 28, 29]. In addition, engineered B. subtilis has been used for the production of industrial biopolymers such as poly‑γ‑glutamic acid, further highlighting its potential as a microbial cell factory [30]. However, engineering B. subtilis for HA production presents specific genetic and metabolic challenges. Unlike natural HA producing organisms such as Streptococcus species, B. subtilis lacks a native hyaluronan synthase gene responsible for the polymerization of HA from UDP sugar precursors. Therefore, reconstruction of the HA biosynthetic pathway needs heterologous expression of a functional hyaluronan synthase and B. subtilis only carries precursor-related genes such as tuaD (UDP-glucose dehydrogenase) [21, 22, 31]. In addition, efficient HA synthesis depends on a balanced intracellular supply of key precursors. Limitations in precursor availability and metabolic flux distribution can restrict HA yield, making pathway optimization and targeted metabolic engineering essential for achieving efficient HA production in this host [22, 24]. In addition to heterologous expression of HA synthase, efficient HA production in B. subtilis needs precise metabolic rewiring. This involves redirecting the carbon flux by modulating the competing pathways such as glycolysis and the pentose phosphate pathway while maintaining cellular energy balance. For example, complete inactivation of the pfkA gene is lethal, indicating that these pathways must be carefully fine-tuned rather than fully disrupted [22]. Recent developments in strain engineering and bioprocess optimization with advances in enzyme/pathway engineering have enabled the identification of improved UDP‑sugar pathway enzymes and hyaluronan synthases that enhance HA biosynthesis in non‑pathogenic hosts [21, 22, 32–34].
In this study, a B. subtilis strain was developed that was capable of efficient HA production through heterologous expression of the hyaluronan synthase gene, which is absent in the native genome and NX02_04625, a functional homolog of tuaD that enhances the supply of UDP-GlcUA precursors [22, 24]. Single and dual-gene expression systems were also constructed and compared. By introducing a novel genetic combination of hyaluronan synthase gene with an additional tuaD homolog from another bacterial source with favorable functional traits, this approach establishes a novel expression platform that addresses biosafety concerns associated with pathogenic production strains and provides a promising foundation for scalable HA biosynthesis. Unlike previous studies that overexpressed the native tuaD or used various hyaluronan synthase sources, this study co‑expressed a codon‑optimized hyaluronan synthase from S. dysgalactiae with a high‑performance tuaD homolog (NX02_04625) from Sphingomonas sanxanigenens, a combination not previously reported in B. subtilis. This approach potentially improves critical metabolic difficulties identified in previous studies and contributes to the advancement of safe and efficient microbial platforms for HA production.
- Materials and Methods
- Bacterial strains and growth conditions
Escherichia coli (E. coli) DH5α ATCC 53868 used for plasmid propagation and B. subtilis 168 ATCC 23857 used as the expression host for HA production were provided by the National Institute of Genetic Engineering and Biotechnology. Cultures were grown in Luria-Bertani (LB) medium (10 g l⁻¹ peptone, Merck, Germany; 5 g l⁻¹ yeast extract, Merck, Germany; 10 g l⁻¹ NaCl, Merck, Germany). Precultures were prepared by inoculating a single colony into 5 ml of LB medium and incubating overnight at 37 °C with agitation at 180 rpm. For plasmid maintenance, appropriate antibiotics were added to the medium, including ampicillin (Sigma‑Aldrich, USA) at a final concentration of 100 μg ml⁻¹ for E. coli and chloramphenicol (Sigma‑Aldrich, USA) at a final concentration of 10 μg ml⁻¹ for B. subtilis. For production experiments, the overnight preculture was transferred into fresh medium at 1% (v/v). The selected production medium contained sucrose (30 g l⁻¹; Merck, Germany), yeast extract (10 g l⁻¹; Merck, Germany), soybean peptone (5 g l⁻¹; Merck, Germany), ammonium sulfate (1 g l⁻¹; Merck, Germany), K₂HPO₄ (9.15 g l⁻¹; Merck, Germany), KH₂PO₄ (3 g l⁻¹; Merck, Germany) and trisodium citrate (1 g l⁻¹; Merck, Germany) [22].
Gene expression was induced by adding isopropyl β‑D‑1‑thiogalactopyranoside (IPTG) to a final concentration of 1 mM (Thermo Fisher Scientific, USA) at the beginning of cultivation and the cultures were incubated at 30 °C with agitation at 180 rpm. Samples were collected at designated time points (24, 48 and 72 h after inoculation of the production culture, depending on the experiment) and centrifuged at 2400× g for 5 min to collect cell‑free supernatants, which were used for further analyses.
- Gene selection and vector design
To confer HA biosynthetic capability to B. subtilis 168, hyaluronan synthase gene from S. dysgalactiae ATCC12394 was selected based on its catalytic performance reported in the Braunschweig Enzyme Database. The gene encodes a hyaluronan synthase used for construct assembly and experimental procedures. To further enhance HA production, NX02_04625 from S. sanxanigenens DSM19645 was coexpressed, which was a tuaD homolog with favorable kinetic characteristics.
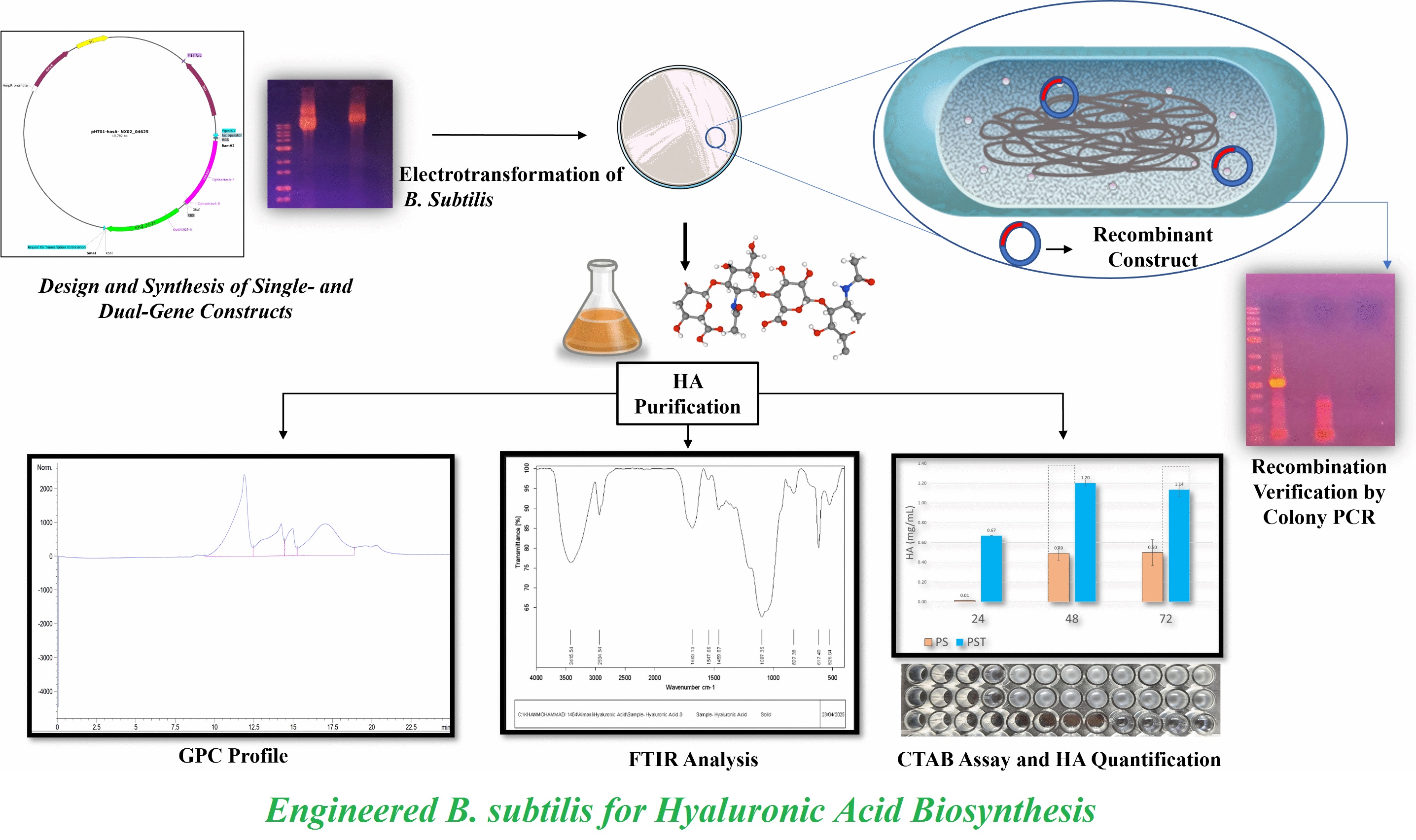
The two genes were codon-optimized for B. subtilis (designed using SnapGene v.3.2.1; GSL Biotech, USA). These were then inserted into the shuttle vector pHT01 under the IPTG-inducible promoter Pgrac01, forming a single transcriptional unit with an intergenic ribosome binding site to enable independent translation. A schematic representation of the dual-gene construct (pHT01-hyaluronan synthase gene-NX02_04625), designed for HA production in B. subtilis 168 (Figure 1), synthesized and sequence-verified by Gene Universal, USA, through its official representative of Pars Biotek, Iran. A single-gene control plasmid was constructed in house.
2.3. Cloning procedures and construct validation
The dual-gene plasmid, hereafter referred to as PST, was reconstituted and diluted to a working concentration. Chemically competent E. coli DH5α cells were transformed using the standard heat-shock method with minor modifications. Transformed cells were plated on LB agar containing ampicillin [35]. To excise the NX02_04625 region, the PST was digested with the XbaI restriction enzyme (Thermo Fisher Scientific, USA; lot no. 01258845), resolved on agarose gel and the PS was excised and purified using FavorPrep gel/PCR purification mini kit (Favorgen Biotech, Taiwan; cat. no. FAGCK001) according to the manufacturer’s instructions. The purified backbone-hyaluronan synthase gene, referred to as PS, was ligated using T4 DNA ligase (Thermo Fisher Scientific, USA; lot no. 00024630).
Ligation products were transformed into E. coli DH5α and plated on ampicillin LB agar. Plasmid DNA was extracted using plasmid DNA extraction mini kit (Favorgen Biotech, Taiwan; cat. no. FAPDE050) according to the manufacturer’s instructions. Construct sizes were verified using gel electrophoresis.
To prepare 0.7% (w/v) agarose gels, 0.7 g of agarose was dissolved in 100 ml of 0.7× tris-borate-EDTA (TBE) buffer. The mixture was heated using microwave. After cooling down, the gel was poured into a casting tray and a comb was inserted to form the wells. After solidification, the gel was transferred into an electrophoresis tank filled with 0.7× TBE buffer. The DNA samples were loaded and separated at a constant voltage of 5–8 V cm⁻¹ until adequate migration was achieved. Band sizes were determined by comparison with 1-kb plus DNA ladder (SinaClon, Iran; cat. no. SL7052).
- Transformation into Bacillus subtilis 168 and validation
The minimum inhibitory concentration (MIC) of chloramphenicol for B. subtilis 168 was assessed as 10 μg ml⁻¹. Electrocompetent B. subtilis cells were transformed with PS and PST constructs via electroporation [36] and selected on LB-chloramphenicol plates. Colonies were validated using colony PCR and gene-specific primers synthesized by Genfanavaran, Iran. Colony PCR was carried out on B. subtilis transformants to verify insertion of recombinant constructs.
- Hyaluronic acid purification
From the media [27, 37], only the modified Westbrook formulation [22] supported HA synthesis. Culture supernatants from engineered B. subtilis were clarified by centrifugation and subjected to sequential trichloroacetic acid (Merck, Germany) and ethanol (Merck, Germany) precipitation. For trichloroacetic acid precipitation, 100 µl of 100% (w/v) trichloroacetic acid were added to 1 ml of clarified supernatant, followed by incubation on ice for 30 min and centrifugation at 16,200× g for 15 min at 4 °C. The resulting supernatant was mixed with 2 ml of cold ethanol (absolute), transferred to -70 °C for 2 h and centrifuged at 4,400× g for 20 min at 4 °C. The pellet was washed with 1 ml of cold ethanol (absolute), incubated at -20 °C for 1 h and recentrifuged. After air‑drying, the HA pellet was dissolved in 1 ml of deionized water and incubated at 45 °C with agitation until fully solubilized. Samples were stored at 4 °C until further analysis.
- Cetyltrimethylammonium bromide turbidimetric method
The HA levels were quantified using cetyltrimethyl-ammonium bromide turbidimetric assay. Samples were mixed with acetate buffer (0.2 M, pH 6; Merck, Germany) and cetyltrimethylammonium bromide reagent (2.5% w/v in 0.5 M NaOH; Merck, Germany) and absorbance was measured at 540 nm. A standard curve prepared from commercial HA (Sigma-Aldrich, USA; lot no. BCBN4845V) was used for the quantification, ensuring a consistent linear relationship between absorbance and HA concentration within the assay working range [38].
- Fourier-transform infrared analysis
Fourier-transform infrared (FTIR) spectroscopy was used to verify the structural identity of the purified HA. Spectra were recorded in the range of 4000–400 cm⁻¹ using standard scanning parameters on the Bruker FTIR spectrometer (Bruker, USA). Characteristic peaks corresponding to hydroxyl, carboxyl and amide groups were assessed.
- Molecular weight determination by size‑exclusion chromatography
Size‑exclusion chromatography separates molecules based on their hydrodynamic size, which depends on molecular weight and molecular conformation in solution. This technique is historically known as gel filtration or gel permeation chromatography (GPC) [39]. The GPC was carried out to assess molecular weight distribution of the purified HA samples. The system (Waters, USA) was equipped with two PL aquagel-OH 8 µm mixed-H columns (300 × 7.5 mm each, separation range 6,000–10⁷ g mol⁻¹; Agilent Technologies, USA) and a refractive index detector (Waters 2414, Waters, USA). Deionized water was used as the mobile phase at a flow rate of 1.0 ml min⁻¹ and the column temperature was set at 35 °C. Calibration was carried out using polyethylene glycol standards (Agilent Technologies, USA), ranging from 1.03 × 10³ to 1.18 × 10⁶ g mol⁻¹. All samples were filtered through 0.45-µm membrane filters (Merck Millipore, USA) prior to injection.
- Statistical analysis
All experiments were carried out with at least two biological replicates and results were expressed as mean ±SD (standard deviation). Normality of data was assessed graphically. Statistical comparisons were carried out using Microsoft Excel v.16.79, USA, and GraphPad Prism v.9.5.0, USA. The HA concentrations, calculated based on optical density values and validated standard curve, were used for statistical analyses. For comparison between the wild-type and PST strains, Welch’s two‑tailed t‑test was used to account for unequal variances. One‑tailed t‑test was used to compare HA production between the PS and PST strains. Paired t‑test was used for temporal comparisons (24 against 48 h, 48 against 72 h) within each strain. A p‑value less than 0.05 was considered statistically significant.
- Results and Discussion
3.1. Construction of the single-gene construct
Polymerase chain reaction analysis of the PST verified the expected amplicon sizes for the two target regions, validating construct integrity. Transformation into E. coli DH5α yielded robust colony formation on selective medium and plasmid extraction produced DNA of the expected size, verifying plasmid stability. Digestion with XbaI generated fragments of the anticipated sizes, enabling recovery and successful assembly of the PS construct.
3.2. Verification of single-gene construct via plasmid size comparison
Size analysis of the plasmid preparations showed a clear distinction between the two constructs. Figure 2 illustrates the agarose gel electrophoresis of the PS and PST constructs. The upward shift observed for the PST variant verified the expected size increase, consistent with the presence of NX02_04625 in PST and its successful excision during generation of the PS construct.
3.3. Introduction of single and dual-gene constructs into Bacillus subtilis 168
The PS and PST constructs were successfully established in B. subtilis 168, as evidenced by the formation of discrete and stable colonies on selective medium. Colony-level analysis verified the expected inserts in B. subtilis 168 transformants. The PS and PST transformants respectively produced amplicons, indicating the presence of PS and PST constructs and verifying successful uptake and stable maintenance in the host strain.
- Initial assessment of hyaluronic acid yield in wild-type against dual-gene recombinant Bacillus subtilis
The comparison between the wild‑type and PST‑harboring B. subtilis 168 strains in LB medium revealed clear evidence of HA production. Figure 3 illustrates the HA concentrations produced by these two strains at 24 and 48 h. The PST recombinant produced 0.22 ±0.03 and 0.31 ±0.02 g l⁻¹ HA at 24 and 48 h respectively, whereas only a negligible signal was observed in the wild-type strain, likely due to background turbidity in the assay. The differences between the two strains were statistically significant at the two sampling times, indicating the HA biosynthetic capacity conferred by the PST construct.
- Comparison of hyaluronic acid production in single and dual-gene-engineered Bacillus subtilis strains
Modified Westbrook fermentation medium was used for comparative evaluation of the PS and PST strains [22]. Figure 4 shows the HA concentrations produced by PS and PST at 24, 48 and 72 h post-induction. In the PST, HA levels reached 1.20 ±0.03 g l⁻¹ at 48 h and 1.14 ±0.07 g l⁻¹ at 72 h with no significant difference between the two time points (p > 0.05). The PS produced 0.49 ±0.07 g l⁻¹ at 48 h and 0.50 ±0.13 g l⁻¹ at 72 h, showing no significant temporal change (p > 0.05). At the two sampling times, HA production in the PST strain was higher than that in the PS strain (p = 0.064), suggesting improved biosynthetic performance of the dual‑gene construct.
The PST construct demonstrated an advantage over the PS construct, achieving a HA yield of 1.20 ±0.03 g l⁻¹ in shake-flasks compared to 0.50 ±0.13 g l⁻¹, a 2.4-fold increase. This clear difference highlighted critical difficulties in the precursor supply chain, specifically the conversion of UDP-glucose to UDP-GlcUA, which apparently improved by the expression of NX02_04625. This finding aligned with and reinforced the metabolic engineering principle established by Westbrook et al. [22], who identified the increase of the UDP-GlcUA pool via the native tuaD gene as the most impactful intervention, boosting the HA titer 5.6‑fold in their B. subtilis system.
A broader comparison of HA yield and molecular weight reported in previous studies provided useful context for interpreting the current results. The B. subtilis systems produce approximately 1–2 g l⁻¹ HA in shake-flask cultures. Under fed-batch fermentation conditions, Corynebacterium glutamicum achieved up to 28.7 g l⁻¹ (0.21 MDa) [40] and 21.6 g l⁻¹ (1.28 MDa) [41]. Studies on B. subtilis in shake-flask systems have reported yields nearly 1.2 g l⁻¹ [22, 24], with molecular weights ranging from 1.5–2.0 MDa [22]. Zhang et al. reported a yield of 1.8 g l⁻¹ under similar conditions [37]. It is noteworthy that the reported HA yields depended strongly on cultivation conditions; fed-batch bioreactor systems generally achieve higher titers than shake-flask cultures. Therefore, the values should be interpreted as approximate comparisons between various production platforms, rather than direct performance equivalents.
The core of our engineering strategy was the rational reconstruction and enhancement of the heterologous HA biosynthetic pathway, enabling a final HA yield of 1.20 g l⁻¹ in shake-flasks. Native B. subtilis possesses the genetic machinery for UDP-N-acetylglucosamine (UDP-GlcNAc) synthesis but lacks the hyaluronan synthase gene and shows limited flux toward the second precursor of UDP-GlcUA [22, 31]. The present approach involved the heterologous coexpression of hyaluronan synthase gene from S. dysgalactiae and a high-performance tuaD homolog (NX02_04625) from S. sanxanigenens, selected for its favorable enzymatic kinetics from the Braunschweig Enzyme Database.
- Fourier-transform infrared analysis of produced hyaluronic acid
Figure 5 shows the FTIR spectrum of purified HA produced by engineered B. subtilis 168 carrying the PST plasmid. This spectrum showed the characteristic absorption bands of HA, including O–H stretch (3415 cm⁻¹), C–H stretch (2934 cm⁻¹), asymmetric and symmetric COO⁻ stretches (1683 and 1459 cm⁻¹) and C–O–C vibration at 1097 cm⁻¹. The current characterization methods supported the biosynthesis of HA. The FTIR spectroscopy showed absorption bands attributable to key functional groups (O–H, COO⁻ and C–O–C) consistent with previously reported HA spectra [42]. This was consistent with functional heterologous expression of the hyaluronan synthase gene, a critical step toward biomedical applicability [43].
- Molecular weight distribution by size‑exclusion chromatography
Figure 6 illustrates results of size‑exclusion chromatography analysis of purified HA produced by engineered B. subtilis 168 carrying the PST plasmid. This revealed a trimodal distribution.
- Molecular weight parameters
Quantitative MW data for fractions within the calibration range are reported in Table 1. The HMW-HA (Peak 1) exceeded the column calibration range; its MW was estimated as > 1.18 × 10⁶ g mol-¹ but needed absolute quantification. The GPC analysis revealed a heterogeneous molecular weight distribution, which was typical for microbial HA production due to the presence of endogenous hydrolases or shear stress [44, 45]. The trimodal distribution indicated significant heterogeneity. The dominant HMW fraction (43.45%) aligned with native HA intrinsic chain length, while the mid and LMW fractions (56.55% combined) suggested partial hydrolysis during processing. The narrow PDI (1.20) of fragmented HA suggested controlled degradation, whereas the broader PDI (1.34) of oligomers reflected heterogeneous chain cleavage. In the present study, the engineered B. subtilis strain produced HA with a dominant molecular weight exceeding 1.18 MDa. The HMW-HA (>1.18 MDa) is highly valued in medical and food uses [46, 47]. While this heterogeneity might need further purification for specific uses, it presented an opportunity as low and medium-MW-HA fractions include their own unique therapeutic and food grade uses [48–50]. The average molecular weight of the main fragmented fraction was calculated at 9.88 × 105 g mol-¹, which was within the highly desirable range for commercial uses [51].
A previous study reported that HMW-HA increased the hardness of ginkgo seed protein isolate gels and improved in vitro digestion by up to 70%. Gel texture and digestibility could be controlled by adjusting pH and HA molecular weight, likely due to physical entanglement between HA and protein molecules that restricted their movement and stabilized the gel network [52]. The HMW-HA enhanced texture (via stronger gel networks) and nutrition‑related attributed such as digestibility. Its mucoadhesive characteristics prolonged the contact time of substances with taste receptors, aiding in the perception of saltiness [50].
The escalating demand for HA in the food, pharmaceutical, biomedical and cosmetic industries necessitates a paradigm shift away from traditional, limited production methods. While extraction from animal tissues increases concerns about pathogen contamination and batch-to-batch variability and fermentation using pathogenic Streptococcus strains introduces significant biosafety and regulatory hurdles, the development of efficient GRAS microbial platforms has become a central focus of metabolic engineering. The advantage of the current B. subtilis-based system includes its combination of a GRAS status, high secretory capacity, well-known genetics and absence of endotoxins, which may facilitate safer development and regulatory consideration for food and biomedical uses, compared to E. coli [53].
In summary, this study provides a minimalistic, effective genetic engineering strategy for HA production in B. subtilis. By co-expressing hyaluronan synthase gene and a high-performance tuaD homolog, the study helped decrease precursor bottlenecks under the tested conditions, achieving shake-flask titers that are competitive with early non-pathogenic systems. The GRAS status, high secretory capacity and well-characterized genetics of B. subtilis suggest that this platform may represent a safer alternative to conventional animal-derived or pathogenic fermentation systems. The findings are similar to the authors’ recent complementary studies on HA production systems. In those studies, other HA-producing strains were assessed under various conditions and the regulatory mechanisms of HA biosynthesis were investigated using genome-scale approaches [54, 55]. Insights from those studies contributed to the design and development of the engineered strain in the present study.
Although the present study was carried out at the shake‑flask scale, scaling up HA production to bioreactor systems may introduce several engineering and physiological challenges. Accumulation of HA can increase broth viscosity; thereby, limiting mixing efficiency and oxygen transfer. The HA biosynthesis competes with cellular metabolism for key precursors such as UDP-GlcNAc and UDP-GlcUA, potentially imposing metabolic burden during high‑density cultivation. Furthermore, agitation and aeration affect HA production and molecular weight, while excessive shear stress may damage cells and decrease polymer size. Accumulation of byproducts such as lactic acid may inhibit cell growth and HA synthesis, highlighting the need of careful bioprocess design during scale‑up [19]. The high viscosity of HA-rich fermentation broths poses significant challenges for downstream processing, including filtration and recovery of HA. These challenges necessitate careful optimization of the purification steps, especially during scale-up to industrial production [56].
- Conclusion
In this study, the dual-gene-engineered strain demonstrated enhanced HA yield, compared to the single-gene-engineered strain, highlighting the critical role of precursor supply in HA biosynthesis. Characterization of the purified HA by FTIR verified its chemical identity, showing all expected functional groups, while GPC analysis revealed a predominantly HMW polymer fraction, suitable for food, biomedical and cosmetic uses. The findings establish this engineered B. subtilis platform for HA biosynthesis, with the potential to meet growing market demand for safe and high quality HA. This study highlights B. subtilis as a promising chassis for the safe and efficient biomanufacturing of high-value biopolymers.
In addition to the scientific and technical contributions, this study includes broader industrial significance. The establishment of a GRAS, non‑pathogenic and secretion‑competent B. subtilis platform provides a further safe, regulatory‑friendly cost‑effective alternative to animal‑derived or pathogenic fermentation routes currently used in the biopharmaceutical and cosmetic industries. The ability to produce HMW-HA, a premium grade material for dermal fillers, wound healing products, moisturizers and functional foods, highlights the translational potential of this platform as a promising foundation for further industrial bioprocess development. However, translating this proof‑of‑concept into an industrially relevant platform needs further optimization of process scale-up, molecular weight control and plasmid stability.
Based on the limitations identified in this study, further research should focus on improving HA production in B. subtilis through metabolic engineering and process optimization. Although a titer of 1.20 g l-¹ was achieved at shake-flask scale without optimization, additional studies are needed to enhance production efficiency and assess plasmid stability during scale-up. Further studies investigate hydrolase gene deletion to improve molecular weight, optimization of fed-batch fermentation and engineering strategies to decouple production from sporulation for further scalable and stable processes.
- Declaration
5.1. Acknowledgements
This study was carried out as a part of the first author's PhD project.
5.2. Declaration of competing interest
The authors declare no conflict of interest.
5.3. Authors’ Contributions
Rouzbeh Almasi Ghale: Conceptualization; data curation; formal analysis; investigation; methodology; validation; visualization; writing – original draft; writing – review & editing.
Reza Faghihi: Investigation; formal analysis; writing – original draft; visualization.
Marjan Talebi: Formal analysis; writing – original draft; writing – review & editing; visualization.
Mehdi Shamsara: Conceptualization; formal analysis; investigation; methodology; supervision; validation; writing – original draft; writing – review & editing.
Fatemeh Tabandeh: Conceptualization; formal analysis; investigation; methodology; project administration; funding acquisition; supervision; validation; writing – original draft; writing – review & editing.
5.4. Using Artificial Intelligent Chatbots
The authors used ChatGPT (OpenAI) for text refinement and improving readability of the manuscript. All scientific content, data interpretation and final responsibility for the manuscript are linked to the authors.
5.5. Ethical Consideration
None.
- Bacillus subtilis 168
- Hyaluronan synthase gene
- Hyaluronic acid production
- Metabolic engineering
- Strain development
- UDP glucose dehydrogenase
How to Cite
References
1. Wu Y, Zhao S, Wang J, Chen Y, Li H, Li JP, Kan Y, Zhang T. Methods for determining the structure and physicochemical properties of hyaluronic acid and its derivatives: A review. Int J Biol Macromol. 2024; 282: 137603. https://doi.org/10.1016/j.ijbiomac.2024.137603
2. Saadati F, Bahrulolum H, Talebi M, Karimi M, Bozorgchami N, Almasi Ghale R, Zafar S, Aghighi Y, Asiaei E, Tabandeh F. Advances and principles of hyaluronic acid production, extraction, purification and its applications: A review. Int J Biol Macromol. 2025; 312: 143839. https://doi.org/10.1016/j.ijbiomac.2025.143839
3. Garantziotis S, Savani RC. Hyaluronan biology: A complex balancing act of structure, function, location and context. Matrix Biol. 2019; 78: 1-10. https://doi.org/10.1016/j.matbio.2019.02.002
4. Bukhari SNA, Roswandi NL, Waqas M, Habib H, Hussain F, Khan S, Sohail M, Ramli NA, Thu HE, Hussain Z. Hyaluronic acid, a promising skin rejuvenating biomedicine: A review of recent updates and pre-clinical and clinical investigations on cosmetic and nutricosmetic effects. Int J Biol Macromol. 2018; 120: 1682-1695. https://doi.org/10.1016/j.ijbiomac.2018.09.188
5. Vasvani S, Kulkarni P, Rawtani D. Hyaluronic acid: A review on its biology, aspects of drug delivery, route of administrations and a special emphasis on its approved marketed products and recent clinical studies. Int J Biol Macromol. 2020; 151: 1012-1029. https://doi.org/10.1016/j.ijbiomac.2019.11.066
6. Liu M, Tolg C, Turley E. Dissecting the dual nature of hyaluronan in the tumor microenvironment. Front Immunol. 2019; 10: 947. https://doi.org/10.3389/fimmu.2019.00947
7. Silva AL, Babo PS, Rodrigues MT, Gonçalves AI, Novoa-Carballal R, Pires RA, Rouwkema J, Reis RL, Gomes ME. Hyaluronic acid oligomer immobilization as an angiogenic trigger for the neovascularization of TE constructs. ACS Appl Bio Mater. 2021; 4(8): 6023-6035. https://doi.org/10.1021/acsabm.1c00291
8. Jiang Y, Jiang Y, Li L, Liu X, Hou X, Wang W. High-molecular-weight hyaluronic acid can be used as a food additive to improve the symptoms of persistent inflammation, immunosuppression and catabolism syndrome (PICS). Biology (Basel). 2024; 13(5): 319. https://doi.org/10.3390/biology13050319
9. Zhou HD, Tang SH, Li SN, Liao BX, Ma Y, Luo ZL, Wang JX. Effect of adding hyaluronic acid with various molecular masses on the quality of yogurt. Preprint. 2023:189-197. https://doi.org/10.7506/spkx1002-6630-20230105-028
10. Huang X, Chen Z, Liu D, Zheng Z, Li Y, Zhu Y, Yan ZY, Hu J, Zhou P. Concentration and molecular weight of hyaluronic acid contributes to sweet taste perception. SSRN Electron J. [Preprint] 2023. Available from: https://dx.doi.org/10.2139/ssrn.4504005
11. Guo L, Wang P, Jiang Z, Guo Y, Zhou F, Chai Y. Effect of hyaluronic acid coating on fresh quality of crucian carp (Carassius auratus) during the partial-freezing storage. Food Mach. 2017; 33(12):120-125. https://doi.org/10.13652/j.issn.1003-5788.2017.12.024
12. Razzak MA, Kim M, Kim HJ, Park YC, Chung D. Deciphering the interactions of fish gelatine and hyaluronic acid in aqueous solutions. Int J Biol Macromol. 2017; 102: 885-892. https://doi.org/10.1016/j.ijbiomac.2017.04.083
13. Koe T. Exclusive to Korea: Cenovis targets females in 20s with new hyaluronic acid drink. NutraIngredients-Asia. Available from: https://www.nutraingredients-asia.com/Article/2020/12/23/Exclusive-to-Korea-Cenovis-targets-females-in-20s-with-new-hyaluronic-acid-drink/ [Accessed 28 May 2026].
14. Sola R, Valls RM, Martorell I, Giralt M, Pedret A, Taltavull N, Romeu M, Rodriguez A, Morina D, De Frutos VL. A low-fat yoghurt supplemented with a rooster comb extract on muscle joint function in adults with mild knee pain: a randomized, double blind, parallel, placebo-controlled, clinical trial of efficacy. Food Funct. 2015; 6(11): 3531-3539. https://doi.org/10.1039/C5FO00321K
15. Cheng Q, Liu C, Zhao J, Li W, Guo F, Qin J, Wang Y. Unlocking the potential of hyaluronic acid: Exploring its physicochemical properties, modification and role in food applications. Trends Food Sci Technol. 2023; 142: 104218. https://doi.org/10.1016/j.tifs.2023.104218
16. Al-Hilifi SA, Al-Ali RM, Dinh LN, Yao Y, Agarwal V. Development of hyaluronic acid based polysaccharide-protein composite edible coatings for preservation of strawberry fruit. Int J Biol Macromol. 2024; 259: 128932. https://doi.org/10.1016/j.ijbiomac.2023.128932
17. Fan Z, Hao Y, Wang Y, Hu X, Li T. Characterisation of hyaluronic acid-curcumin-cellulose nanofibre composite film and application in egg preservation. Int J Food Sci Technol. 2023; 58(12): 6263-6271. https://doi.org/10.1111/ijfs.16729
18. Shikina E, Kovalevsky R, Shirkovskaya A, Toukach PV. Prospective bacterial and fungal sources of hyaluronic acid: A review. Comput Struct Biotechnol J. 2022;20: 6214-6236. https://doi.org/10.1016/j.csbj.2022.11.013
19. Liu L, Liu Y, Li J, Du G, Chen J. Microbial production of hyaluronic acid: current state, challenges and perspectives. Microb Cell Fact. 2011; 10(1): 99. https://doi.org/10.1186/1475-2859-10-99
20. Grabowski M, Gmyrek D, Żurawska M, Trusek A. Hyaluronic acid: production strategies, gel-forming properties and advances in drug delivery systems. Gels. 2025; 11(6): 424. https://doi.org/10.3390/gels11060424
21. Manfrão-Netto JH, Queiroz EB, Oliveira Junqueira AC, Gomes AM, Gusmão de Morais D, Paes HC, Parachin NS. Genetic strategies for improving hyaluronic acid production in recombinant bacterial culture. J Appl Microbiol. 2022; 132(2): 822-840. https://doi.org/10.1111/jam.15242
22. Westbrook AW, Ren X, Oh J, Moo-Young M, Chou CP. Metabolic engineering to enhance heterologous production of hyaluronic acid in Bacillus subtilis. Metab Eng. 2018; 47: 401-413. https://doi.org/10.1016/j.ymben.2018.04.016
23. Su Y, Liu C, Fang H, Zhang D. Bacillus subtilis: a universal cell factory for industry, agriculture, biomaterials and medicine. Microb Cell Fact. 2020; 19(1):173. https://doi.org/10.1186/s12934-020-01436-8
24. Widner B, Behr R, Von Dollen S, Tang M, Heu T, Sloma A, Sternberg D, DeAngelis PL, Weigel PH, Brown S. Hyaluronic acid production in Bacillus subtilis. Appl Environ Microbiol. 2005; 71(7): 3747-3752. https://doi.org/10.1128/AEM.71.7.3747-3752.2005
25. Zhao X, Chen X, Xue Y, Wang X. Development of an efficient iterative genome editing method in Bacillus subtilis using the CRISPR-AsCpf1 system. J Basic Microbiol. 2022; 62(7): 824-832. https://doi.org/10.1002/jobm.202200134
26. Xiang M, Kang Q, Zhang D. Advances on systems metabolic engineering of Bacillus subtilis as a chassis cell. Synth Syst Biotechnol. 2020; 5(4): 245-251. https://doi.org/10.1016/j.synbio.2020.07.005
27. Jin P, Kang Z, Yuan P, Du G, Chen J. Production of specific-molecular-weight hyaluronan by metabolically engineered Bacillus subtilis 168. Metab Eng. 2016; 35: 21-30. https://doi.org/10.1016/j.ymben.2016.01.008
28. Liu Y, Zhang Q, Qi X, Gao H, Wang M, Guan H, Yu B. Metabolic engineering of Bacillus subtilis for riboflavin production: a review. Microorganisms. 2023; 11(1):164. https://doi.org/10.3390/microorganisms11010164
29. Liu ZY, Yu XZ. Engineering Bacillus subtilis for high-value bioproduction: recent advances and applications. Microb Cell Fact. 2025; 24(1): 182. https://doi.org/10.1186/s12934-025-02818-6
30. Tian Y, Hu R, Zhang R, Yan G, Zhang Y, Hu J, Zhao S, Ju J, Yu B, Wang L. Engineering of Bacillus subtilis KH2 for effective production of poly-γ-glutamic acid from glutamate waste based on transcriptomics. Int J Biol Macromol. 2025; 149708. https://doi.org/10.1016/j.ijbiomac.2025.149708
31. Jia Y, Zhu J, Chen X, Tang D, Su D, Yao W, Gao X. Metabolic engineering of Bacillus subtilis for the efficient biosynthesis of uniform hyaluronic acid with controlled molecular weights. Bioresour Technol. 2013; 132: 427-431. https://doi.org/10.1016/j.biortech.2012.12.150
32. Marwan-Abdelbaset E, Samy-Kamal M, Tan D, Lu X. Microbial production of hyaluronic acid: The current advances, engineering strategies and trends. J Biotechnol. 2025; 403: 52-72. https://doi.org/10.1016/j.jbiotec.2025.03.015
33. Yao ZY, Yu MJ, Li QQ, Gong JS, Zhang P, Jiang JY, Su C, Xu G, Jia BY, Xu ZH. Unlocking green biomanufacturing potential: superior heterologous gene expression with a T7 integration overexpression system in Bacillus subtilis. ACS Synth Biol. 2024; 14(6): 1977-1987. https://doi.org/10.1021/acssynbio.4c00694
34. Cerminati S, Leroux M, Anselmi P, Peirú S, Alonso JC, Priem B, Menzella HG. Low cost and sustainable hyaluronic acid production in a manufacturing platform based on Bacillus subtilis 3NA strain. Appl Microbiol Biotechnol. 2021; 105(8): 3075-3086. https://doi.org/10.1007/s00253-021-11506-5
35. Froger A, Hall JE. Transformation of plasmid DNA into E. coli using the heat shock method. J Vis Exp. 2007; (6):e253. https://doi.org/10.3791/253
36. Harwood CR, Cutting SM. Molecular biological methods for Bacillus. Chichester: Wiley; 1990.
37. Chien LJ, Lee CK. Enhanced hyaluronic acid production in Bacillus subtilis by coexpressing bacterial hemoglobin. Biotechnol Prog. 2007; 23(5): 1017-1022. https://doi.org/10.1021/bp070036w
38. Chen YH, Wang Q. Establishment of CTAB turbidimetric method to determine hyaluronic acid content in fermentation broth. Carbohydr Polym. 2009; 78(1): 178-181. https://doi.org/10.1016/j.carbpol.2009.04.037
39. López-Cánovas AE, Victoria-Sanes M, Martínez-Hernández GB, López-Gómez A. Methods for determining the high molecular weight of hyaluronic acid: A review. Polymers (Basel). 2025; 17(24): 3289. https://doi.org/10.3390/polym17243289
40. Cheng F, Yu H, Stephanopoulos G. Engineering Corynebacterium glutamicum for high-titer biosynthesis of hyaluronic acid. Metab Eng. 2019; 55: 276-289. https://doi.org/10.1016/j.ymben.2019.07.003
41. Cheng F, Luozhong S, Guo Z, Yu H, Stephanopoulos G. Enhanced biosynthesis of hyaluronic acid using engineered Corynebacterium glutamicum via metabolic pathway regulation. Biotechnol J. 2017; 12(10): 1700191. https://doi.org/10.1002/biot.201700191
42. Fan D, Wang B, Zheng X, Guo Q. Determination of hyaluronan by spectroscopic methods. J Wuhan Univ Technol Mater Sci Ed. 2006; 21(3): 32-34. https://doi.org/10.1007/BF02840874
43. Fallacara A, Baldini E, Manfredini S, Vertuani S. Hyaluronic acid in the third millennium. Polymers (Basel). 2018; 10(7): 701. https://doi.org/10.3390/polym10070701
44. Gomes AMV, Manfrão-Netto JH, Carvalho LS, Parachin NS. Heterologous hyaluronic acid production in Kluyveromyces lactis. Microorganisms. 2019; 7(9):294. https://doi.org/10.3390/microorganisms7090294
45. Güngör G, Gedikli S, Toptaş Y, Akgün DE, Demirbilek M, Yazıhan N, Aytar Çelik P, Denkbaş EB, Çabuk A. Bacterial hyaluronic acid production through an alternative extraction method and its characterization. J Chem Technol Biotechnol. 2019; 94(6):1843-1852. https://doi.org/10.1002/jctb.5957
46. Kogan G, Šoltés L, Stern R, Gemeiner P. Hyaluronic acid: a natural biopolymer with a broad range of biomedical and industrial applications. Biotechnol Lett. 2007; 29(1): 17-25. https://doi.org/10.1007/s10529-006-9219-z
47. Tabasi A, Tabandeh F, Maghsoudi A, Bafrouei SMM, Khodabandeh M, Ghasemi F, Ranjbar M. Improved production of food-grade hyaluronic acid in recombinant Corynebacterium glutamicum by medium optimization and feeding strategy. Appl Food Biotechnol. 2022; 12(1):1-14. http://dx.doi.org/10.22037/afb.v12i1.46588
48. Tammi RH, Passi AG, Rilla K, Karousou E, Vigetti D, Makkonen K, Tammi MI. Transcriptional and post-translational regulation of hyaluronan synthesis. FEBS J. 2011; 278(9): 1419-1428. https://doi.org/10.1111/j.1742-4658.2011.08070.x
49. Talebi M, Almasi Ghale R, Mokhtari Asl R, Tabandeh F. Advancements in characterization and preclinical applications of hyaluronic acid-based biomaterials for wound healing: A review. Carbohydr Polym Technol Appl. 2025; 100706. https://doi.org/10.1016/j.carpta.2025.100706
50. Sutariya SG, Joshi R, Salunke P. Emerging applications of hyaluronic acid in dairy and foods. Discov Food. 2026; 6:84. https://doi.org/10.1007/s44187-025-00776-x
51. La Gatta A, Schiraldi C, Papa A, De Rosa M. Comparative analysis of commercial dermal fillers based on crosslinked hyaluronan: Physical characterization and in vitro enzymatic degradation. Polym Degrad Stab. 2011; 96(4):630-636. https://doi.org/10.1016/j.polymdegradstab.2010.12.025
52. Cheng Q, Liu C, Zhao J, Guo F, Qin J, Wang Y. Hyaluronic acid modulates techno-functional and digestion properties of heat-induced ginkgo seed protein isolate gel. Food Biosci. 2024; 59:104204. https://doi.org/10.1016/j.fbio.2024.104204
53. van Dijl J, Hecker M. Bacillus subtilis: from soil bacterium to super-secreting cell factory. Microb Cell Fact. 2013; 12(1):3. https://doi.org/10.1186/1475-2859-12-3
54. Nikuiyan Z, Tabandeh F, Motamedian E, Salehi S, Talebi M, Almasi Ghale R. Reconstruction of a genome-scale metabolic model for Streptococcus zooepidemicus: Comparison with Corynebacterium glutamicum to study hyaluronic acid production. PLoS One. 2025; 20(12):e0335509. https://doi.org/10.1371/journal.pone.0335509
55. Nadali Hazaveh M, Talebi M, Almasi Ghale R, Zilouei H, Tabandeh F. Engineered biosynthesis of hyaluronic acid in Corynebacterium glutamicum and green synthesis of HA-silver nanocomposites for advanced antimicrobial wound dressings. Sci Rep. 2026; 16:7910. https://doi.org/10.1038/s41598-026-39148-5
56. Flores-Gatica M, Castañeda-Aponte H, Gil-Garzon MR, Mora-Galvez LM, Banda-Magaña MP, Jáuregui-Jáuregui JA, Torres-Acosta MA, Mayolo-Deloisa K, Licona-Cassani C. Primary recovery of hyaluronic acid produced in Streptococcus equi subsp. zooepidemicus using PEG–citrate aqueous two-phase systems. AMB Express. 2021; 11(1):123. https://doi.org/10.1186/s13568-021-01287-5
- Abstract Viewed: 0 times
- pdf Downloaded: 0 times