
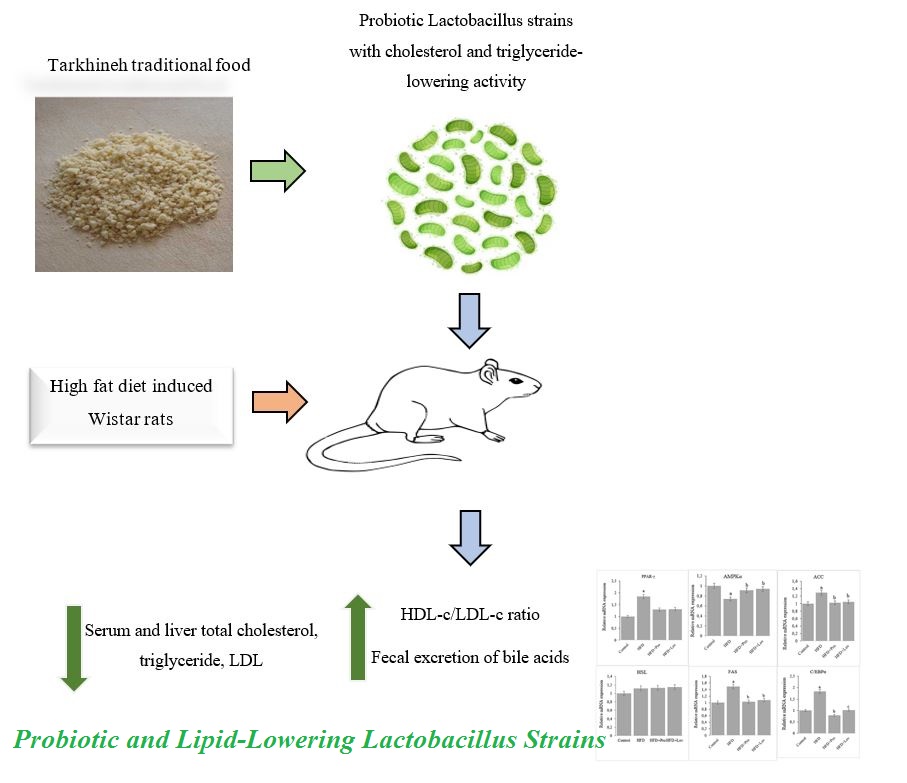
Antihyperlipidemic and Probiotic Characteristics of Novel Lactobacillus Strains Isolated from Traditional Iranian Dairy
التكنولوجيا الحيوية الغذائية التطبيقية,
مجلد 12 عدد 1 (2025),
4 كانون الثاني 2025
,
الصفحة 1-13 (e31)
https://doi.org/10.22037/afb.v12i1.50629
الملخص
Background and Objective: This study aimed to investigate the antihyperlipidemic effects of potent probiotic Lactobacillus strains isolated from traditional Iranian dairy in a high-fat diet rat model.
Material and Methods: Lactobacillus strains were isolated from tarkhineh samples and screened for significant in-vitro cholesterol and triglyceride-decreasing activities, with key probiotic characteristics. Seven strains were selected based on their high in-vitro lipid-decreasing activity, substantial resistance to simulated gastric and intestinal conditions, resistance to 0.5% phenol and 15 mg l-1 lysozyme, adhesion capacity to Caco-2 cells, antibiotic susceptibility profiles, and antagonistic activity against human pathogens in male Wistar rats fed a high-fat diet. The lipid-decreasing activity was assessed in male Wistar rats (n = 7 per group) over 6 weeks, with the probiotic mixture administered daily at a dose of 2 × 109 CFU ml-1 rat-1.
Results and Conclusion: Using 16S rDNA analysis, these strains were identified as Lactobacillus casei, Lactobacillus fermentum, Lactobacillus kefiri, Lactobacillus alimentarius, Lactobacillus acidophilus, Lactobacillus reuteri, and Lactobacillus brevis. The lipid-decreasing activity was assessed in male Wistar rats. Compared to the high-fat diet control group, the probiotic-supplemented diet decreased serum total cholesterol by 16.6% and LDL-cholesterol by 56.7% (p < 0.05). Particularly, the probiotic mixture resulted in a 13.2-fold increase in the HDL-c/LDL-c ratio (from 0.132 to 1.75), compared to high-fat diet controls (p < 0.01). Mechanistically, the probiotic diet increased fecal cholic acid excretion by 4.7-fold (from 1.17 to 5.54 µmol g-1) (p < 0.05) and decreased hepatic steatosis. Treatment attenuated high-fat diet-induced upregulation of lipogenic genes (PPAR-γ, ACC, FAS, and C/EBPα) and restored the expression of AMPKα. These results indicated that supplementation with lactobacilli from homemade dairy was effective in improving dyslipidemia, suggesting these products could be a promising source of novel probiotics.
Keywords: Probiotics, Traditional dairy, Lactobacillus spp., Tarkhineh, Hyperlipidemia
- Introduction
High levels of serum lipids are significant risk factors for cardiovascular disease (CVD). Individuals with dyslipidemia have a threefold higher risk of heart attack, compared to those with normal lipid levels. This condition is often related to dietary habits, particularly the consumption of unhealthy diets. Diets high in fat, especially saturated fatty acids (SFA), can increase blood total cholesterol (TC) and triglyceride (TG) levels, which increases the risk of atherosclerosis, coronary heart disease, and stroke [1, 2].
Medications such as statins and fibric acid derivatives are commonly used to manage blood lipid levels. However, these conventional drugs can include adverse effects, including muscle pain, liver damage, neurological disorders, increased blood sugar, and miscarriage. Therefore, there is an increasing need for safer and further cost-effective dietary interventions to address imbalanced serum lipids. Probiotics have emerged as a potential solution for managing dyslipidemia, and their use in functional foods is a promising area of research. Probiotics have been shown to significantly assimilate cholesterol and TG from culture media. Bacteria such as Lactobacillus, Lactococcus, and Bifidobacterium spp. are commonly detected in fermented dairy products [3-5].
Traditional fermented foods contain unique microbial communities that are shaped by local production methods and environmental conditions [6]. Within the lactic acid bacteria (LAB) family, Lactobacillus spp. are the most common members. These bacteria are often isolated from various Iranian traditional fermented foods. One such food is tarkhineh, a dried fermented mixture of yoghurt, cracked wheat, and vegetables, which offers a stable acidic matrix conducive to LAB preservation. The dry acidic nature of tarkhineh allows for the long-term preservation of its milk proteins [7].
While numerous studies have screened probiotics for cholesterol-decreasing characteristics, most isolates originate from well-characterized sources such as commercial yogurts or dairy products with standardized fermentation [8]. The wide microbial diversity of traditional, regionally distinct Iranian fermented foods, such as tarkhineh, as a source of novel strains with potentially superior or unique functionalities, is largely uninvestigated. Moreover, tarkhineh offers a unique ecological niche as a stable acidic matrix with high solid contents. This environment imposes selective pressure on the microbiota, favoring highly robust and acid-resistant Lactobacillus strains [7, 9].
These environmental stressors suggest that isolates from tarkhineh may possess superior functional traits such as enhanced resistance to gastric conditions and greater bile-salt hydrolase activity, compared to strains from conventional sources [10]. Furthermore, studies of novel isolates depend solely on in vitro screening, failing to provide the necessary comprehensive in vivo validation. The precise molecular mechanisms underlying the antihyper-lipidemic effects, particularly regarding the modulation of key hepatic lipogenic genes, are often not fully clarified for novel isolates. Therefore, this study aimed to fill this critical gap by providing a full characterization of novel Lactobacillus strains from tarkhineh, followed by a robust, mechanistic in vivo validation of their efficacy in a high-fat diet (HFD) rat model.
- Materials and Methods
2.1 Isolation and Preliminary Identification of Lactobacilli
Samples of tarkhineh (n = 22) were collected during July and August 2024 from various rural areas of Lorestan Province, Iran, including Dowlatabad and Mahrouw Villages in Aligoudarz County (33°09′N 49°24′E) and Dehnow and Emamabad Villages in Dorud County (33°29′58″N 49°03′11″E). To prevent microbial contamination and changes to the primary microbiota, samples were prepared under hygienic conditions and stored at 4 °C. For isolation, 5 g of each sample was mixed with 50 ml of sodium citrate (Merck, Germany) and homogenized using a stomacher. The samples were serially diluted up to 10⁻⁷ in sterile saline solution and cultured on de Man, Rogosa, and Sharpe (MRS) agar (HiMedia, India). After anaerobic incubation (Gas-pack system) at 37 °C for 48 h, colonies were inoculated into 10 ml of MRS broth and incubated at 37 °C for 24 h. The isolates were screened through primary morphological assessment using a fluorescent microscope (Zeiss Axioskop, Germany) and standard biochemical assessments, including catalase, oxidase, and carbohydrate fermentation. Screened lactobacilli were preserved at -70 °C in glycerol and skim milk.
2.2 Bile Salts and Low pH Tolerance
The ability of isolates to tolerate bile salts was assessed as described by Saboori et al. [11]. Each strain (2% v/v) was grown in MRS broth containing 0.3% (w/v) bile salts (0% as a control). Cultures were incubated at 37 °C for 6 h, and optical density (OD) was measured at 600 nm. For acid tolerance, strains were cultured overnight in MRS broth adjusted to pH values of 2.0, 2.5, and 6.4 (as a control). The OD values at 600 nm were recorded at the beginning and end of cultivation. Tolerance was calculated using methods described by Liu et al. [12].
2.3 In-vitro Cholesterol and Triglyceride Decreasing Activities
The cholesterol-decreasing ability of strains was assessed using the method by Liu et al. [12]. Strains were grown in MRS broth at 37 °C for 24 h and then inoculated into MRS broth supplemented with cholesterol and 0.3% bile salts (MRS-CHOL). A control sample of uninoculated MRS-CHOL was also prepared. After 24 h of incubation at 37 °C without shaking, the bacterial broth was centrifuged, and cholesterol content in the cell-free supernatant was assessed. Strains that lowered cholesterol by more than 50% were selected for triglyceride (TG) decreasing assessments. These strains were grown overnight in MRS broth and then transferred to MRS-TG broth. This was prepared by mixing 20 ml of 2% polyvinyl alcohol solution with 50 ml of triglycerides (Merck, Germany) and adding the mixture to MRS broth (3% v/v). The pH was adjusted to 6.5, and the medium was sterilized. After 72 h of incubation at 37 °C, the TG quantity in the cell-free supernatant was assessed using a commercial kit (Cayman, USA) [13].
2.4 Cell Survival in Simulated Gastrointestinal Juice
To screen for tolerance to gastrointestinal stress, isolated lactobacilli were cultured in MRS broth and incubated anaerobically overnight at 37 °C. The suspension was centrifuged at 6000× g for 7 min, and the cells were washed twice with sterile phosphate-buffered saline (PBS, pH 2.0). Aliquots were cultured on MRS agar and incubated anaerobically at 37 °C for 0, 1, 2, and 3 h. The number of viable bacteria was assessed as colony-forming units per milliliter (CFU ml-1). For assessment under simulated conditions, overnight cultures of isolates were centrifuged at 6000× g for 5 min, washed with 50 mM PBS (pH 6.5), and dissolved in 3 ml of PBS buffer. One-ml aliquots of each isolate (containing 9-log CFU ml-1 of bacteria) were mixed with 9 ml of simulated gastric juice (7 mM KCl, 45 mM NaHCO₃, 125 mM NaCl, and 3 g l-1 pepsin at pH 2.5). The mixtures were incubated at 37 °C for 3 h and then centrifuged at 4000× g for 7 min. The pellets were washed three times with PBS and resuspended in simulated intestinal juice (pH 8.0) containing 0.15% (w/v) bile salt and 0.1% (w/v) pancreatin. Then, the suspensions were incubated at 37 °C for another 3 h, and the viable bacteria were counted and reported as Log CFU ml-1.
2.5 Phenol and Lysozyme Tolerance
Phenol tolerance is critical for lactobacilli survival in the gastrointestinal tract (GIT), as phenol can be produced by gut microbiota. Bacterial cultures grown for 24 h were cultured in MRS broth containing 0.5% phenol (Sigma, USA). The ODs were measured at 580 nm using a microplate reader (Thermo Fisher Scientific, USA). Lysozyme tolerance was assessed as described by Zafar et al. [14]. Bacterial cells were collected (4000× g, 5 min), washed, and resuspended in PBS. Then, 10 µl of cell suspensions were transferred to PBS solutions containing 15 mg l-1 lysozyme (0% as a control). After 2 h of incubation at 37 °C, ODs were measured at 600 nm to estimate survival rates.
2.6 Safety Assessment
2.6.1 Antibiotic Susceptibility
The antibiotic susceptibility of isolated lactobacilli was assessed based on the method of Saboktakin et al. [15]. Strains (1.5 × 10⁸ CFU ml-1) were cultured on MRS agar. Antibiotic discs were transferred onto the surface, set at room temperature (RT) for 10 min for diffusion, and then incubated at 37 °C overnight. The areas of inhibition around each disc were assessed.
2.6.2 DNase and Hemolytic Activities
To assess DNase activity, isolates were streaked on DNase agar (HiMedia, India) and incubated at 37 °C for 72 h, and then clear zones were assessed. Staphylococcus aureus was used as a positive control. Hemolytic activity was assessed on blood agar containing 5% v/v sheep blood. Isolates were cultured on the media and incubated at 37 °C for 48 h. Blood lysis zones were characterized as α-hemolysis (green zones), β-hemolysis (clear zones), or γ-hemolysis (no zones). Only isolates showing γ-hemolysis were considered safe [16].
2.7 Antibacterial Activity Assessment
Antibacterial spectra were assessed using agar diffusion assay [17]. The indicator bacteria were five human pathogens of Salmonella typhimurium ATCC 14028, Escherichia coli ATCC 25922, Listeria monocytogenes CMCC 54002, Bacillus subtilis ATCC 11060, and Staphylococcus aureus ATCC 25923. These pathogens were overlaid on Mueller-Hinton (M-H) agar plates with 7-mm diameter wells. Freshly prepared cell-free supernatants of each isolate were filtered (0.2-µm filter), poured into the wells, and incubated at 37 °C overnight. Then, diameters of the inhibitory zones were assessed.
2.8 Assessment of the Isolates' Adhesion Ability
To assess the ability of strains to adhere to Caco-2 cells, an in vitro model for intestinal epithelia was assessed using the method of Greene and Klaenhammer [18]. Caco-2 cells were grown in Dulbecco's Modified Eagle Medium (DMEM) (Sigma, USA) supplemented with 1% penicillin-streptomycin and fetal bovine serum (FBS) in a 5% CO₂ atmosphere at 37 °C for 20 days. Cells were transferred to tissue culture plates and grown to a monolayer. The plates were washed with sterile PBS to remove the media, particularly penicillin-streptomycin. Overnight cultures of bacterial strains were centrifuged (5000 rpm, 5 min), washed, and resuspended in DMEM to a concentration of 10⁸ bacteria ml-1. This suspension was added to Caco-2 monolayer cells and incubated at 37 °C for 1 h. Unbound bacteria were removed by washing with PBS. Then, Caco-2 cells were lysed with Triton X-100 (0.1% v/v), and bacterial counts were carried out on MRS agar. Adhesion capacity was calculated using the Eq 1:
Adhesion capacity (%) = (A / B) × 100 Eq.1
Where A was the number of attached bacteria, and B was the total number of bacteria added to each well.
2.9 Molecular Identification and Phylogenetic Analysis
Genomic DNA was extracted from overnight bacterial cultures using a commercial kit (Pars Azmoun, Iran). A conserved region of the 16S rDNA gene was amplified using 27F-1492R primers as described by Nami et al. Amplicons were electrophoresed on a 2% agarose gel. The amplified fragments were purified using the PureLink PCR purification kit (Thermo Fisher Scientific, USA) and sequenced by BGI Biotechnology (Shenzhen, China). Sequences were compared to the GenBank database via BLAST similarity search; isolates with over 98% similarity to reference sequences were assigned to the same species. A phylogenetic tree was constructed using MEGA7 software (Biodesign Institute, USA) with the Kimura 2-parameter model and the neighbor-joining method [19].
2.10 In vivo Study
A total of 24 male Wistar rats (age, 6 weeks; weight, 150 g ±15) were purchased from the Pasteur Institute, Tehran, Iran. All procedures were carried out based on the ARRIVE guidelines and approved by the ethical committee of Islamic Azad University (IR.IAU.ARAK.REC.1399.021).
Experiments were based on the guidelines in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health (NIH, USA). Rats were acclimated for 1 week with commercial foods (Specialty Feeds, USA) and water to minimize environmental stress. The animals were housed in a controlled environment (23 °C, 55% relative humidity, 12-h light/dark cycle). The rats were randomly allocated into four groups of six animals, with similar initial body weights. The groups received the following diets for 35 days:
- ND, Normal diet
- HFD, High-fat diet
- HFD and Pro, HFD with a potential probiotic mixture (2 × 10⁹ CFU ml-1 d-1)
- HFD and Lov, HFD with lovastatin (15 mg kg-1 d-1)
The HFD was composed of 78.3% commercial diet, 5% lard oil, 5% corn germ oil, 5% sucrose, 5% dried egg yolk, 1% cholesterol, 0.5% sodium deoxycholate, and 0.2% propylthiouracil. All administrations were carried out via intragastric gavage. The inclusion of propylthiouracil was a deliberate methodological choice to establish a robust model of complex, resistant dyslipidemia. The probiotic dose was selected based on prior studies demonstrating efficacy in rodent models [12, 20]. Blood samples were collected under ketamine-xylazine anesthesia on Days 0 and 35. Serum was extracted, and TC, high-density lipoprotein (HDL), low-density lipoprotein (LDL), and TG levels were assessed using commercial kits (Pars Azmoun, Iran). At the end of the experiment, rats were euthanized using Forane (isoflurane). Livers were extracted for histological studies using hematoxylin and eosin (H&E) staining and for lipid analysis using a method described by Folch et al. Feces were collected on Days 33, 34, and 35 for cholic acid analysis.
2.11 Real-time Polymerase Chain Reaction
The expression levels of six lipid metabolism-linked genes were assessed using quantitative real-time PCR (qRT-PCR) [21]. Primers were generated for peroxisome proliferator-activated receptor-γ (PPAR-γ), adenosine 5′-monophosphate-activated protein kinase-α (AMPKα), CAATT/enhancer-binding protein α (C/EBPα), hormone-sensitive lipase (HSL), fatty acid synthetase (FAS), acetyl-CoA carboxylase (ACC), and the internal control GAPDH (Table 1). Total RNA was extracted from tissues using TRIzol reagent (Invitrogen, USA), purified with a Qiagen RNeasy mini kit (QIAGEN, Germany), and quantified by spectrometry (Eppendorf, Germany). The cDNA was synthesized using the RevertAid first-strand cDNA synthesis kit (Thermo Fisher Scientific, USA). The qRT-PCR reactions were carried out with SYBR Premix Ex Taq II kit (Takara, China) on a PRIMEPRO48 real-time qPCR thermal cycler (Antylia Scientific, USA). Results were calculated using the 2_ΔΔCT method and are shown as fold changes.
2.12 Statistical Analysis
Data were analyzed using SPSS software v.18 (IBM, USA) through a one-way analysis of variance (ANOVA). Differences between the means were reported using the Duncan test at a 95% confidence level (p < 0.05). All experiments were carried out in triplicate.
- Results and Discussion
3.1 Isolation and Screening of Potential Probiotic Lactobacilli
A total of 83 Gram-positive, catalase-negative, coccoid bacterial isolates were collected from tarkhineh samples, all of which showed a carbohydrate fermentation profile characteristic of the Lactobacillus genus. These isolates were used in a bile-salt tolerance prescreening, from which 36 strains showing an inhibition rate below 50% were selected for acid tolerance assessment. Of these, 19 isolates demonstrated survival rates higher than 85% at pH values of 2.5 and 2.0.
High levels of cholesterol and triglycerides in the diet and serum are major risk factors for cardiovascular and metabolic diseases. Since pharmacotherapy can cause adverse effects, it is important to find natural and non-toxic substances to decrease TC, LDL, and TG levels. Lactobacilli, which have been used in fermented dairy products for many years and are generally considered safe (GRAS) [22], have been shown to effectively decrease lipids in animal and clinical research [23, 24]. In this study, seven potential probiotic lactobacilli strains were isolated from Iranian traditional dairy products. These strains showed enhanced cholesterol and triglyceride-decreasing activity, increased tolerance to simulated gastric and intestinal juice, improved adhesion to human intestinal cells, and expanded antibacterial spectrum under in vitro conditions.
3.2 In-vitro Cholesterol and Triglyceride Assimilations
Of the 19 acid-tolerant strains, seven strains showed cholesterol-decreasing rates exceeding 50%. Of these, three strains demonstrated triglyceride (TG)-decreasing rates of more than 40%. As shown in Table 2, the highest cholesterol-decreasing rate (67.41% ±0.20) was observed for Strain L-42, while the highest TG-decreasing rate (51.15% ±0.57) belonged to Strain L-3.
In vitro, the L-42 and L-11 strains showed cholesterol-decreasing rates of 67.41 and 64.27%, respectively. These rates were higher than those of L. fermentum F1 (48.87%) [25], L. acidophilus L2-16 (64.11%) [26], Pediococcus acidilactici LAB 5 (62%) [27], L. sake C2 (53.2%) [28], B. longum (34.2%) [29] and L. helveticus MG2-1 (51.74%) [28].
There are a few investigations on the ability of potential probiotics in decreasing TG. The rate for the L-3 strain was higher than that L. acidophilus L2-73 reported in the Gao et al. study [26]. The authors observed that cholesterol and TG intake varied greatly in strains within the same species. This suggested that cholesterol sequestering was specific to individual strains rather than a characteristic of the entire species [30].
3.3 Resistance to Simulated Gastrointestinal Conditions, Phenol, and Lysozyme
All seven selected strains, except L-51 (69% survival), showed a survival rate above 80% after sequential exposure to simulated gastrointestinal fluids. The strains were highly resistant to simulated gastric fluid, with over 94.8% survival after 2 h and over 90% survival for most strains after 4 h. In simulated intestinal fluid, six isolates survived above 90%. Overall, the strains were relatively more tolerant to acid than bile. There was no significant difference between strains in their tolerance to simulated fluids, which was similar to the initial pH and bile salt assessments. All bacteria tolerated 0.5% phenol after overnight incubation (OD > 1.000) and showed significant resistance to 15 mg l-1 lysozyme after 2 h of exposure (Table 3).
Probiotics must be able to tolerate gastrointestinal stressors such as acidity, bile, phenol, lysozyme, and pepsin. The current research showed that lactobacilli isolated from fermented dairy demonsterated strong tolerance to simulated gastrointestinal environments.
3.4 Safety Profile of Antibiotic Resistance, DNase, and Hemolytic Activity
The isolated lactobacilli showed various resistance to the assessed antibiotics (Table 4). Chloramphenicol, novobiocin, and penicillin were effective against all the isolates. In contrast, most isolates were resistant to rifampicin. All seven lactobacilli were negative for DNase activity and showed no hemolytic activity (γ-hemolysis) on blood agar, indicating that they were safe.
3.5 Antagonistic Activity against Pathogens
Six of the seven lactobacilli showed moderate antipathogenic activities; strain L-42 did not demonstrate antagonistic activity. The metabolites from the isolates were generally more effective against Gram-negative pathogens such as S. typhimurium and E. coli than Gram-positive ones such as B. subtilis and L. monocytogenes (Table 5).
The isolated strains showed strong antagonistic activity against Gram-positive and Gram-negative pathogens. This is a key beneficial effect, as it helps maintain a balanced intestinal flora in the host and manage gastrointestinal infections.
3.6 Adhesion to Caco-2 Cells
The adhesion rates of the lactobacilli to Caco-2 cells varied between the strains of L-3 (24.2% ±0.11), L-11 (13.3% ±0.11), L-20 (28.9% ±0.05), L-24 (10.8% ±0.24), L-42 (11.5% ±0.21), L-48 (10.6% ±0.42), and L-51 (31.8% ±0.37). Strains L-3, L-20, and L-51 demonstrated significantly higher adhesion capabilities compared to the other strains (p < 0.05). Probiotics must show antimicrobial activity against intestinal pathogens and adhere to the intestinal epithelia. The current in vitro screening showed significant functional differences between the seven Lactobacillus strains, typical of natural isolates. This diversity justified using mixed culture in vivo to target hyperlipidemia through multiple mechanisms. Strain L-42 was the top cholesterol-decreasing agent (67.41%) with strong metabolic activity but no pathogen inhibition, making it a purely metabolic contributor. In contrast, Strains L-24 and L-48 included moderate lipid-decreasing effects but excelled in gastric tolerance, cell adhesion, and pathogen antagonism, supporting gut ecology and safety. This strain-specific variation highlighted a functional consortium, where each strain played unique roles. The in vivo success was likely a result of synergy between Strain L-42 metabolic effects and the survival and protective traits of the other strains.
3.7 Molecular Identification and Phylogenetic Analysis
All seven isolates showed more than 98% 16S rRNA sequence similarity with Lactobacillus spp. The phylogenetic tree (Figure 1) showed that Strains L-3, L-11, L-20, L-24, L-42, L-48, and L-51 clustered with L. casei, L. fermentum, L. kefiri, L. brevis, L. acidophilus, L. reuteri, and L. alimentarius, respectively.
The biochemical characteristics of these strains were similar to those of lactobacilli, and their nucleotide sequences were homologous to L. casei, L. fermentum, L. kefiri, L. alimentarius, L. acidophilus, L. reuteri, and L. brevis.
3.8 In-vivo Antihyperlipidemic Effects
3.8.1 Effects on Serum Lipid Profile
On Day 0, serum TC, LDL-C, HDL-C, and TG levels were similar in all groups. By Day 35, one-way ANOVA followed by Duncan's post-hoc test verified that the HFD group showed significant increases in serum TC, LDL-C, and TG, while HDL levels significantly decreased by 38.2% (all p < 0.01 compared to the ND group). Compared with the HFD group, HFD and Pro and HFD and Lov groups had significantly lower TC and LDL-C levels and significantly higher HDL levels (all comparisons with p < 0.01, Duncan's test) (Figure 2a). Particularly, the TG level in the HFD and Pro group was significantly lower than that of the HFD group (p < 0.05) and the HFD and Lov group (p < 0.05, Duncan's test). The HDL-c/LDL-c ratio was significantly higher in the HFD and Pro group (1.75 ±0.15), compared to the ND (p < 0.01), HFD (p < 0.01), and HFD and Lov (p < 0.05) groups, as assessed by Duncan's post-hoc test. This improvement underscored the probiotics' potential in modifying atherosclerosis risk, though clinical validation in humans is warranted. Comparative effects of normal diet, HFD, probiotic supplementation, and lovastatin on body weight, food intake, and fecal lipid excretion in experimental groups are available in Table 6.
3.8.2 Effects on Liver Lipids and Histology
As shown in Figure 2b, liver TC and TG levels in the HFD group were significantly higher than those in the HFD and Pro and HFD and Lov groups (comparisons with p < 0.01, Duncan's test). The probiotic-treated rats had significantly lower liver TG levels than the lovastatin-treated group (p < 0.05). Histological examination revealed a normal liver structure in the ND group (Figure 2c). In contrast, the HFD group showed a high degree of vacuolization, lipid accumulation, hepatocyte ballooning, inflammatory cell infiltration, and mild necrosis. Liver sections from the HFD and Pro, and HFD and Lov rats showed significantly fewer pathological changes, compared to the HFD group.
The strains were assessed in a rat model to assess their ability to decrease cholesterol and triglyceride levels. Results showed that administering a combination of isolated probiotics successfully decreased serum and liver levels of TC, TG, and LDL-C in Wistar rats fed a high-fat diet, compared to rats fed a similar diet without probiotics or lovastatin supplementation. The findings revealed that probiotics were as effective as lovastatin in improving dyslipidemia. The reduction in serum and liver TC, TG, and LDL by the probiotics mixture was higher than that of L. plantarum YS5 [20], L. plantarum PH04 [31], B. bifidum PRL2010 [30], and E. faecalis L2-73 [26]. The HDL levels and HDL-c/LDL-c ratio increased in the HFD and Pro group, which was similar to the results of several other studies [14, 32, 33]. L. rhamnosus CK102 was a probiotic strain isolated from fermented yogurt. In a mouse model, it decreased TC by 27.9%, HDL by 28.7% and TG by 61.6% [34]. Epidemiological studies have shown a link between CVD and serum TC and TG levels. Similarly, another study detected that L. fermentum MCC2760 decreased TC, TG, and LDL-c levels in mice [35]. In another rat model, L. fermentum PH5 improved blood lipid profiles by decreasing TC (67.21%), TG (66.21%), and LDL-c (63.25%). The L. fermentum PH5 and PD2 decreased liver TC levels, with L. fermentum PH5 performing better than PD2 [36].
3.8.3 Effects on Fecal Cholic Acid Excretion
Fecal cholic acid levels were 5.14 ±0.15 µmol g-1 for the ND group, 1.17 ±0.10 µmol g-1 for the HFD group, 5.54 ±0.18 µmol g-1 for the HFD and Pro group, and 2.16 ±0.19 µmol g-1 for the HFD and Lov group. The HFD and Pro group had significantly higher concentrations of fecal cholic acid than the ND, HFD, and HFD and Lov groups (all comparisons with p < 0.05, Duncan's test). There is increasing evidence that gut microbiota plays a key role in the development of CVD and metabolic disease. The authors detected that rats supplemented with probiotics had increased levels of cholic acids in their feces. This increase in fecal bile acid elimination has been reported in several probiotic strains, including B. bifidum PRL2010 [30] and E. faecalis L2-73 [26]. The current in-vivo study lacked gut microbiota sequencing and key metabolite assessments (SCFAs, BSH). Despite increased fecal bile acid excretion, microbial mechanisms are unverified. Further studies are needed to validate gut-mediated hypolipidemic pathways.
3.8.4 Effects on Hepatic Gene Expression
The HFD caused a significant increase in the mRNA expression of PPAR-γ, ACC, FAS, and C/EBPα, compared to the ND group (all comparisons with p < 0.05, Duncan's test). This upregulation was normalized by supplementation with either the probiotic mixture or lovastatin (Figure 3). The mRNA expression of AMPKα was significantly downregulated in the HFD group (p < 0.01). However, treatment with the probiotic mixture significantly increased the mRNA expression of AMPKα (p < 0.01). The mRNA level of HSL increased, but not significantly.
The authors assessed the expression of lipid metabolism-linked genes in experimental groups to investigate the potential mechanisms by which isolated lactobacilli decreased dyslipidemia induced by a high-fat diet in rats. The PPAR-γ and C/EBP-α are key transcription factors for adipocyte differentiation in various tissues. Previous studies showed that lactobacilli supplementation decreased the expression levels of PPAR-γ and C/EBP-α in high-fat diet-fed animals. In this investigation, probiotics attenuated HFD-induced upregulation of lipogenic genes (PPAR-γ, ACC, FAS, and C/EBPα), with restoration of AMPKα expression. The AMPK pathway regulates lipid metabolism, and the enzyme is a cellular energy sensor that shuts down anabolic pathways such as fatty acid synthesis. The ACC and FAS are key proteins in the fatty acid synthesis process [37]. Activation of AMPK-α stimulates ACC phosphorylation, blocking FAS expression. The HSL is the rate-limiting enzyme in TG breakdown in various tissues. In this study, the expression of AMPK-α was downregulated in the HFD group, compared to the ND group, while ACC and FAS expressions were upregulated. These changes were alleviated by probiotic treatment, similar to the previous studies [21, 38]. Overall, these findings indicated that isolated lactobacilli administration effectively modulated the gut microbiota induced by HFD, improving dyslipidemia-linked indicators.
While the current use of the HFD rat model provided robust evidence of the probiotic mixture efficacy, demonstrating significant normalization of serum lipid profiles, decrease of hepatic steatosis, and mechanistic regulation of key lipogenic genes, it is imperative to acknowledge the inherent limitations of translating these findings directly to human subjects. A primary constraint is linked to the physiological differences between rodents and humans, particularly in lipid metabolism and lipoprotein profiles [39]. Rats are generally HDL-dominant (carrying most cholesterol on HDL particles), whereas humans are LDL-dominant. This fundamental difference can affect the precise way that dietary components and probiotic interventions modulate cholesterol transport and clearance [40].
Additionally, the gut microbiome structure and composition, even in controlled animal settings, may not fully recapitulate the complexity of the human gut environment. Therefore, the promising results from this study served as the foundational preclinical evidence for this specific mixed culture [40]. The necessary step for validating the potential of this Lactobacillus consortium as a functional food for dyslipidemia includes carrying out randomized, double-blind, placebo-controlled clinical trials in hyperlipidemic human populations. These trials must focus on verifying the long-term safety, optimal dosage, and efficacy against human-specific clinical endpoints such as serum LDL-C, non-HDL-C, and atherosclerotic markers.
- Conclusion
In conclusion, homemade traditional dairy was screened for its potential to introduce novel and promising probiotic bacteria due to variable animal feeding and the rare use of antibiotics. Seven potential probiotics were successfully isolated from tarkhineh food, and in vitro assessments showed that several strains showed excellent probiotic potential based on their cholesterol and triglyceride assimilation, survival rate under simulated GIT stress, antibiotic susceptibility, antagonistic activity against pathogens, and adhesion to Caco-2 cells. In vivo assessment showed that feeding animals with isolated probiotics could improve dyslipidemia caused by a high-fat diet. Although the current study included limitations due to the use of Wistar rats and the effects seen in animal models might not be the same in humans, these strains show promise for developing novel probiotic-enriched dairy products targeting dyslipidemia in at-risk populations.
- Acknowledgements
This research was technically supported by Pasargad Laboratory.
- Declaration of competing interest
The authors report no conflict of interest.
- Authors’ Contributions
Designate each author’s contribution using their initials. Conceptualization, P.A and K.A.; methodology, software, validation, formal analysis, H.M.; investigation, resources, data curation, K.A.; writing—original draft preparation, P.A writing review and editing, S.S.; visualization, supervision, and project administration.
- Using Artificial Intelligent Chatbots
No artificial intelligence chatbots have been used in any section of the present study.
- Ethical Consideration
An ethical committee of Islamic Azad University approved the study (IR.IAU.ARAK.REC.1399.021) and experiments were done according to Guide for the Care and Use of Laboratory Animals of National Institutes of Health.
- Probiotics
- Traditional dairy
- Lactobacillus spp.
- Tarkhineh
- Hyperlipidemia
كيفية الاقتباس
المراجع
1. Gadelha CJMU, Bezerra AN. Effects of probiotics on the lipid profile: Systematic review. J Vasc Bras. 2019; 18. https://doi.org/10.1590/1677-5449.190117
2. Oynotkinova O, Nikonov E, Demidova T, Baranov A, Kryukov E, Dedov E, et al. Changes in the intestinal microbiota as a risk factor for dyslipidemia, atherosclerosis and the role of probiotics in their prevention. Ter Arkh. 2020; 92(9): 94-101. https://doi.org/10.26442/00403660.2020.09.000780
3. Sugata M, Kim Y, Rachmah AF, Oei J, Purnama ET, Nitbani ACJ, et al. Assessment of probiotic characteristics of Lactiplantibacillus plantarum SU-KC1a isolated from human breast milk in Indonesia. Appl Food Biotechnol. 2024; 11(1): e31. https://doi.org/10.22037/afb.v11i1.44299
4. Prima HS, Rusfidra R, Yansen F, Maulana F, Gusti MA. Characterization, molecular identification and antimicrobial activity of lactic acid bacteria with potentials as halal probiotics isolated from Rinuak fish (Psilopsis sp.) in Lake Maninjau, West Sumatra, Indonesia. Appl Food Biotechnol. 2025; 12(1): 1-16(e16). https://doi.org/10.22037/afb.v12i1.48866
5. Yadav R, Khan SH, Mada SB, Meena S, Kapila R, Kapila S. Consumption of probiotic Lactobacillus fermentum MTCC: 5898-fermented milk attenuates dyslipidemia, oxidative stress and inflammation in male rats fed on cholesterol-enriched diet. Probiotics Antimicrob Proteins. 2019; 11: 509-518. https://doi.org/10.1007/s12602-018-9447-3
6. Sadrizadeh N, Khezri S, Dehghan P, Mahmoudi R. Antibacterial effect of Teucrium polium essential oil and Lactobacillus casei probiotic on Escherichia coli O157: H7 in Kishk. Appl Food Biotechnol. 2018; 5(3): 131-140. https://doi.org/10.22037/afb.v5i3.21051
7. Mohammadi N, Ostovar N. Chemical composition, fatty acid composition and volatile compounds of a traditional Kurdish fermented cereal food: Tarkhineh. Food Chem Adv. 2023; 2: 100187. https://doi.org/10.1016/j.focha.2023.100187
8. Ishimwe N, Daliri EB, Lee BH, Fang F, Du G. The perspective on cholesterol‐lowering mechanisms of probiotics. Mol Nutr Food Res. 2015; 59(1): 94-105. https://doi.org/10.1002/mnfr.201400245
9. Kiani A, Nami Y, Hedayati S, Jaymand M, Samadian H, Haghshenas B. Tarkhineh as a new microencapsulation matrix improves the quality and sensory characteristics of probiotic Lactococcus lactis KUMS-T18 enriched potato chips. Sci Rep. 2021; 11(1): 12599. https://doi.org/10.1038/s41598-021-91950-8
10. Kiani A, Nami Y, Hedayati S, Elieh Ali Komi D, Goudarzi F, Haghshenas B. Application of Tarkhineh fermented product to produce potato chips with strong probiotic properties, high shelf-life and desirable sensory characteristics. Front Microbiol. 2021; 12: 657579. https://doi.org/10.3389/fmicb.2021.657579
11. Saboori B, Shahidi F, Hedayati S, Javadmanesh A. Investigating the probiotic properties and antimicrobial activity of lactic acid bacteria isolated from an Iranian fermented dairy product, Kashk. Foods. 2022; 11(23): 3904. https://doi.org/10.3390/foods11233904
12. Liu C, Xue W-j, Ding H, An C, Ma S-j, Liu Y. Probiotic potential of Lactobacillus strains isolated from fermented vegetables in Shaanxi, China. Front Microbiol. 2022; 12: 774903. https://doi.org/10.3389/fmicb.2021.774903
13. Rudel LL, Morris M. Determination of cholesterol using o-phthalaldehyde. J Lipid Res. 1973; 14(3): 364-6. https://doi.org/10.1016/s0022-2275(20)33443-7
14. Zafar H, Ain Nu, Alshammari A, Alghamdi S, Raja H, Ali A, et al. Lacticaseibacillus rhamnosus FM9 and Limosilactobacillus fermentum Y57 are as effective as statins at improving blood lipid profile in high cholesterol, high-fat diet model in male Wistar rats. Nutrients. 2022; 14(8): 1654. https://doi.org/10.3390/nu14081654
15. Saboktakin-Rizi M, Alizadeh Behbahani B, Hojjati M, Noshad M. Identification of Lactobacillus plantarum TW29-1 isolated from Iranian fermented cereal-dairy product (Yellow Zabol Kashk): Probiotic characteristics, antimicrobial activity and safety evaluation. J Food Meas Charact. 2021; 15(3): 2615-2624. https://doi.org/10.1007/s11694-021-00850-x
16. Casarotti SN, Carneiro BM, Todorov SD, Nero LA, Rahal P, Penna ALB. In vitro assessment of safety and probiotic potential characteristics of Lactobacillus strains isolated from water buffalo mozzarella cheese. Ann Microbiol. 2017; 67: 289-301. https://doi.org/10.1007/s13213-016-1243-z
17. Yasmin I, Saeed M, Khan WA, Khaliq A, Chughtai MFJ, Iqbal R, et al. In vitro probiotic potential and safety evaluation (hemolytic, cytotoxic activity) of Bifidobacterium strains isolated from raw camel milk. Microorganisms. 2020; 8(3): 354. https://doi.org/10.3390/microorganisms8030354
18. Greene JD, Klaenhammer TR. Factors involved in adherence of lactobacilli to human Caco-2 cells. Appl Environ Microbiol. 1994; 60(12): 4487-94. https://doi.org/10.1128/aem.60.12.4487-4494.1994
19. Zhang W, Yang J, Zhou Z, Deng L, Lai S, Liu H, et al. Screening and evaluation of lactic acid bacteria with probiotic potential from local Holstein raw milk. Front Microbiol. 2022; 13: 2665. https://doi.org/10.3389/fmicb.2022.990267
20. Nami Y, Bakhshayesh RV, Manafi M, Hejazi MA. Hypocholesterolaemic activity of a novel autochthonous potential probiotic Lactobacillus plantarum YS5 isolated from yogurt. LWT. 2019; 111: 876-82. https://doi.org/10.1016/j.lwt.2019.05.071
21. Li H, Liu F, Lu J, Shi J, Guan J, Yan F, et al. Probiotic mixture of Lactobacillus plantarum strains improves lipid metabolism and gut microbiota structure in high fat diet-fed mice. Front Microbiol. 2020; 11: 512. https://doi.org/10.3389/fmicb.2020.00512
22. Rodríguez-Sánchez S, Ramos IM, Seseña S, Poveda JM, Palop ML. Potential of Lactobacillus strains for health-promotion and flavouring of fermented dairy foods. LWT. 2021; 143: 111102. https://doi.org/10.1016/j.lwt.2021.111102
23. Khare A, Gaur S. Cholesterol-lowering effects of Lactobacillus species. Curr Microbiol. 2020; 77(4): 638-44. https://doi.org/10.1007/s00284-020-01899-2
24. Wu Y, Zhang Q, Ren Y, Ruan Z. Effect of probiotic Lactobacillus on lipid profile: A systematic review and meta-analysis of randomized, controlled trials. PLoS One. 2017; 12(6): e0178868. https://doi.org/10.1371/journal.pone.0178868
25. Zeng XQ, Pan DD, Zhou PD. Functional characteristics of Lactobacillus fermentum F1. Curr Microbiol. 2011; 62: 27-31. https://doi.org/10.1007/s00284-010-9710-8
26. Gao Y, Li D. Screening of lactic acid bacteria with cholesterol-lowering and triglyceride-lowering activity in vitro and evaluation of probiotic function. Ann Microbiol. 2018; 68(9): 537-545. https://doi.org/10.1007/s13213-018-1365-5
27. Mandal V, Sen SK, Mandal NC. Effect of prebiotics on bacteriocin production and cholesterol lowering activity of Pediococcus acidilactici LAB 5. World J Microbiol Biotechnol. 2009; 25: 1837-1847. https://doi.org/10.1007/s11274-009-0095-x
28. Bilige M, Liu W, Rina W, Wang L, Sun T, Wang J, et al. Evaluation of potential probiotics properties of the screened Lactobacilli isolated from home-made koumiss in Mongolia. Ann Microbiol. 2009; 59: 493-498. https://doi.org/10.1007/s13213-009-0010-9
29. Shin HS, Park SY, Lee DK, Kim SA, An HM, Kim JR, et al. Hypocholesterolemic effect of sonication-killed Bifidobacterium longum isolated from healthy adult Koreans in high cholesterol fed rats. Arch Pharm Res. 2010; 33: 1425-1431. https://doi.org/10.1007/s12272-010-0917-0
30. Zanotti I, Turroni F, Piemontese A, Mancabelli L, Milani C, Viappiani A, et al. Evidence for cholesterol-lowering activity by Bifidobacterium bifidum PRL2010 through gut microbiota modulation. Appl Microbiol Biotechnol. 2015; 99: 6813-6829. https://doi.org/10.1007/s00253-015-6677-5
31. Nguyen T, Kang J, Lee M. Characterization of Lactobacillus plantarum PH04, a potential probiotic bacterium with cholesterol-lowering effects. Int J Food Microbiol. 2007; 113(3): 358-361. https://doi.org/10.1016/j.ijfoodmicro.2006.08.001
32. Brandão LR, de Brito Alves JL, da Costa WKA, Ferreira GdAH, de Oliveira MP, da Cruz AG, et al. Live and ultrasound-inactivated Lacticaseibacillus casei modulate the intestinal microbiota and improve biochemical and cardiovascular parameters in male rats fed a high-fat diet. Food Funct. 2021; 12(12): 5287-5300. https://doi.org/10.1039/d1fo00465a
33. Miao Z, Zheng H, Liu W-H, Cheng R, Lan H, Sun T, et al. Lacticaseibacillus paracasei K56 attenuates high-fat diet-induced obesity by modulating the gut microbiota in mice. Probiotics Antimicrob Proteins. 2022; 14: 1-12. https://doi.org/10.1007/s12602-022-09950-8
34. Zheng J, Wittouck S, Salvetti E, Franz CM, Harris HM, Mattarelli P, et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901 and union of Lactobacillaceae and Leuconostocaceae. Int J Syst Evol Microbiol. 2020; 70(4): 2782-2858. https://doi.org/10.1099/ijsem.0.004107
35. Kumar M, Nagpal R, Kumar R, Hemalatha R, Verma V, Kumar A, et al. Cholesterol-lowering probiotics as potential biotherapeutics for metabolic diseases. Exp Diabetes Res. 2012; 2012: 905917. https://doi.org/10.1155/2012/905917
36. Thakkar PN, Patel A, Modi HA, Prajapati JB. Hypocholesterolemic effect of potential probiotic Lactobacillus fermentum strains isolated from traditional fermented foods in Wistar rats. Probiotics Antimicrob Proteins. 2020; 12(3): 1002-1011. https://doi.org/10.1007/s12602-019-09605-z
37. Ji A, Chen W, Liu C, Zhang T, Shi R, Wang X, et al. Soy protein compared with whey protein ameliorates insulin resistance by regulating lipid metabolism, AMPK/mTOR pathway and gut microbiota in high-fat diet-fed mice. Food Funct. 2023; 14(1): 412-425. https://doi.org/10.1039/d2fo02636d
38. Qiao Y, Sun J, Xia S, Li L, Li Y, Wang P, et al. Effects of different Lactobacillus reuteri on inflammatory and fat storage in high-fat diet-induced obesity mice model. J Funct Foods. 2015; 14: 424-434. https://doi.org/10.1016/j.jff.2015.02.015
39. Ishikawa M, Saito K, Urata M, Kumagai Y, Maekawa K, Saito Y. Comparison of circulating lipid profiles between fasting humans and three animal species used in preclinical studies: Mice, rats and rabbits. Lipids Health Dis. 2015; 14(1): 104. https://doi.org/10.1186/s12944-015-0105-x
40. Fotschki B, Wiczkowski W, Sawicki T, Sójka M, Myszczyński K, Ognik K, et al. Stimulation of the intestinal microbiota with prebiotics enhances hepatic levels of dietary polyphenolic compounds, lipid metabolism and antioxidant status in healthy rats. Food Res Int. 2022; 160: 111754. https://doi.org/10.1016/j.foodres.2022.111754
- الملخص المشاهدات: 113 الأوقات
- pdf (English) التنزيلات: 20 الأوقات