
Engineering phytase for improved thermostability and catalytic efficiency: Impacts on nutrient bioavailability
Applied Food Biotechnology,
Vol. 12 No. 1 (2025),
4 January 2025
,
Page 1-11(e14)
https://doi.org/10.22037/afb.v12i1.48493
Abstract
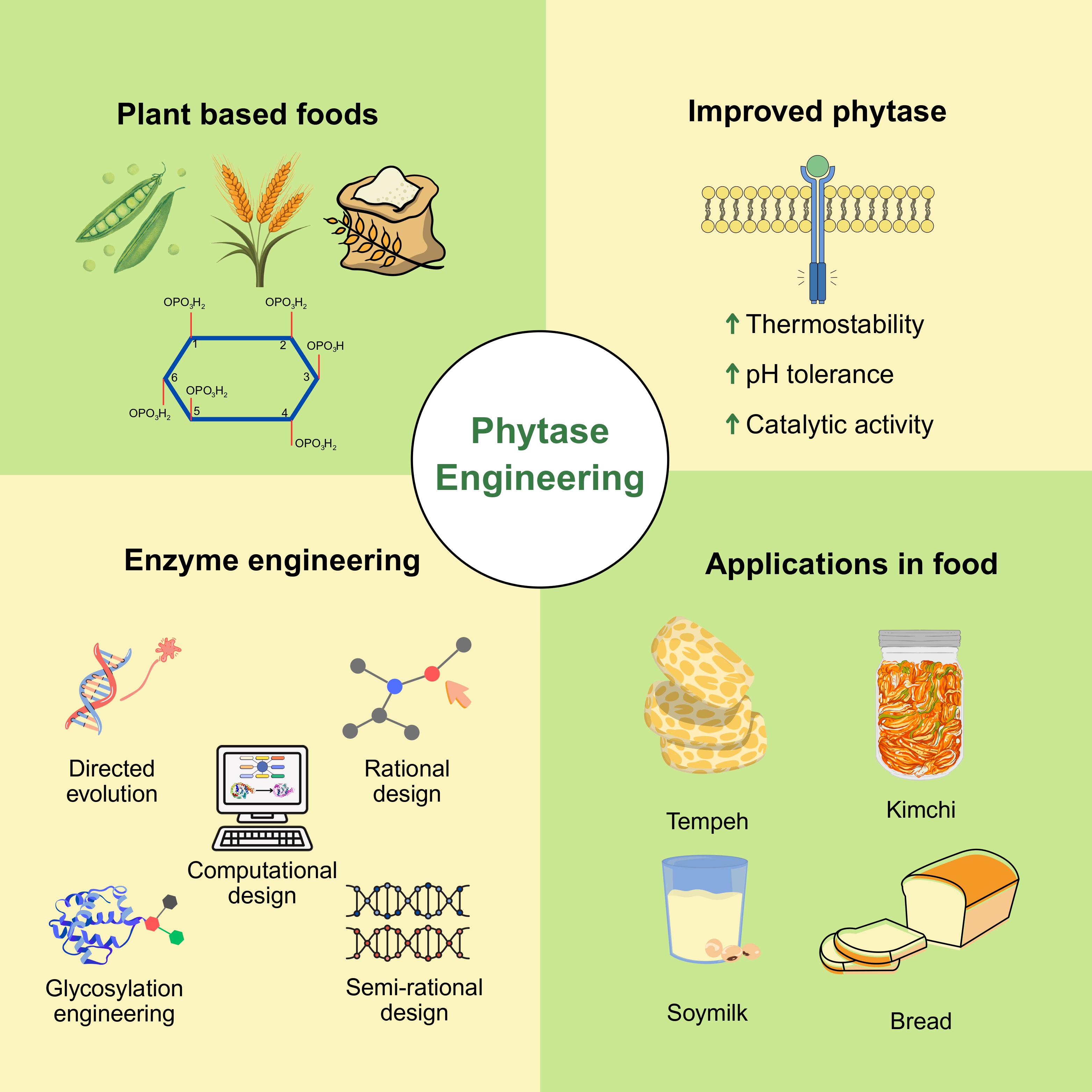
Background and Objective: Phytase is a phosphatase enzyme. It is essential for hydrolyzing phytic acid, an antinutrient present in plant-based foods that chelates essential minerals and limits their bioavailability. Enzyme techniques used to improve phytase characteristics are discussed including thermostability, catalytic efficiency, and substrate specificity.
Results and Conclusion: Phytase efficiency has been significantly improved for industrial applications by different engineering methods, including rational design, computational modeling, directed evolution, glycosylation engineering and semi-rational design. The study further highlights the application of phytase in food processing, including breadmaking, fermented foods, and functional food formulations, to address mineral deficiencies. Future advances in enzyme engineering, such as computational de novo design and enzyme immobilization, could expand commercial and nutritional uses of phytase. The focus of ongoing research on thermostable and highly efficient phytase variants provides significant possibilities for enhancing global food security and sustainability.
Keywords: Phytase, enzyme engineering, directed evolution, food processing, phytic acid, mineral bioavailability
- Introduction
Phytase (EC 3.1.3.8) is also known as myo-inositol hexakisphosphate phosphohydrolase. It is an enzyme that hydrolyzes phytic acid which act as an antinutrient found in plant-based foods. In animals with a single-chambered stomach (monogastric animals like pigs, chickens, and fish), phytate-bound phosphorus is poorly absorbed. In fish, indigestible complexes of phytate and phosphorus form within the digestive tract when they consume phytate-rich plant-based feeds. This leads to reduced phosphorus bioavailability and increased excretion into aquatic environments. Supplementing feed with phytase improves phosphorus availability, supporting enhanced growth, bone development, and overall health while reducing phosphorus waste [1]. Plant-based proteins such as oat, wheat, soy, and pea are widely consumed for their nutritional benefits. They contain significant amount of phytic acid, ranged between 0.4% to 2.2% dry weights depending on the type and processing conditions. These phytate compounds strongly chelate divalent minerals such as calcium, iron, and zinc, significantly reducing their intestinal absorption and contributing to mineral deficiencies, specifically in populations relying heavily on cereals and legumes [2, 5, 6]. It also enhances protein and amino acid digestibility, contributing to better feed efficiency [3].
Phytase is widely applied in the food, feed, and pharmaceutical industries due to its multifunctional properties. The growing demand for stable, high-temperature-resistant phytase has promoted global market growth. In food applications like bread-making, phytic acid limits mineral absorption. Adding phytase during fermentation helps release bound minerals, improving iron and zinc bioavailability. The global phytase market is expected to be 692.5 million USD by 2023, with a compound annual growth rate (CAGR) of 6.3% between 2024 and 2030 [4].
Phytic acid is a major challenge in whole grains and bran, where it chelates minerals and reduces absorption [5]. Based on [7], phytase hydrolyses the phosphate groups attached to the inositol ring to break down phytic acid as shown in Figure 1. Phytase enzymatically hydrolyzes the phosphate ester bonds in phytic acid (myo-inositol hexakisphosphate), sequentially releases inorganic phosphate and lower inositol phosphates. This increases the bioavailability of essential minerals. Phytic acid degradation by phytase supports physiological functions like oxygen transport (via iron) and immune health (via zinc). Enhancing mineral absorption with phytase is important in developing regions with plant-based diets, helping address nutritional deficiencies and improve public health outcomes [6]. However, current enzyme engineering techniques face several limitations. Many phytase variants still lose activity at high processing temperatures or extreme pH conditions. While techniques like directed evolution, rational design, and glycosylation have improved thermostability and activity, they often require extensive trial-and-error, are time-consuming, or lack scalability [17, 24, 36]. Immobilization methods also face challenges in cost-effectiveness and enzyme reuse cycles. Research gaps remain in developing optimum phytase enzymes that retain high activity across a wide range of industrial conditions. There is a need for more in vivo studies validating the nutritional benefits in humans [9].
- Global phytic acid intake from plant-based diet
Phytic acid consumption varies greatly among nations, depending on diet and the volume of plant-based foods consumed. Grains and legumes are majorly consumed in the developing countries, where daily phytic acid consumption can reach more than 2000 mg/day [8]. In developed countries, the daily intake ranges from 250 to 800 mg, depending on their consumption of plant-based diets and whole grains. Phytic acid consumption in China varies from 648 mg to 1433 mg.day-1 across six locations [9]. The average intake was substantially lower among urban people i.e., 781 mg.day-1, compared to rural individuals who consumed approximately 1342 mg.day-1. The amount of phytic acid consumed in the UK increased from 504 to 844 mg.day-1 to 1436 ± 755 mg.day-1 between 1986 and 2005. Children in India between the ages of 4 and 9 consume between 720 and 1160 mg.day-1, while teenagers between the ages of 10 and 19 consume between 1350 and 1780 mg.day-1. Swedish vegetarians consumed 1146 mg.day-1, compared to 180–370 mg.day-1 for those following a Western diet in Finland and Sweden [9]. These variations highlight the influence of dietary patterns on phytic acid consumption, with higher intakes observed in populations that depended heavily on plant-based diets.
Recent studies highlight how dietary habits and food processing influence phytic acid intake and its nutritional impact. It was found that germination and fermentation of cereals and legumes reduced phytic acid content by 50–70% [10]. This significantly improved iron and zinc absorption in plant-based diets. The traditional African processing methods i.e., soaking and malting, reduced phytic acid levels in maize-based foods and enhanced mineral bioavailability for rural populations [11]. Maize soaked for 24 h found to have reduced phytic acid levels by 20–30%, while malting for 48–72 h decreased phytic acid levels by up to 45% in maize-based foods. These processes improved calcium, iron, and zinc bioavailability for rural Malawian populations. They also found that combining soaking and malting resulted in a synergistic effect and reduced phytic acid by 55–60% in maize porridge. These studies highlighted that while plant-heavy diets significantly increase phytic acid consumption, processing techniques found to have mitigated its anti-nutritional effects.
- Different enzyme engineering methods of phytase
Phytases are widely utilized to improve the bioavailability of phosphorus and other essential minerals in the animal feed industry by breaking down phytic acid in grains. A thermostable phytase can maintain its activity under high temperatures. Phytases primarily act by a nucleophilic attack on the phosphorus atom, facilitated by acidic and basic residues in the active site. Enzyme engineering improved the catalytic properties of the residues as shown in Table 1. Improvements in the stability of the transition state of the substrate during the catalytic cycle result in a high enzyme turnover rate. The addition or optimization of disulfide bonds and salt bridges improves the thermal stability and resistance of phytase during processing. Improving the hydrophobic interactions in the core of the enzyme is also responsible for increased thermal stability. Phytase enzyme engineering is based on several advanced techniques to optimize its characteristics i.e., activity, stability, substrate specificity, and resistance to conditions.
3.1. Directed evolution for thermostable phytase development
Directed evolution replicates natural selection in the lab to develop proteins with desired characteristics. It effectively enhances phytase thermostability, ensuring activity at high temperatures during processes. Random mutations are introduced into the phytase gene by increasing deoxyribonucleic acid (DNA) polymerase error rates during polymerase chain reaction (PCR) (using altered Mg²⁺/Mn²⁺ levels or biased nucleotide analogues) or via chemical mutagens like nitrous acid and ethyl methane sulfonate (EMS). Mutated genes are then cloned into expression vectors and transformed into E. coli for screening [12]. When host cells produce mutant enzymes, a library of phytase enzymes is formed, as shown in Figure 2. The generated library is analyzed for phytase variants with improved properties. Enzyme activity and stability are measured using techniques such as microplate assay and flow cytometry.
Thermostability is determined by exposing the enzymes to high temperatures and measuring any residual activity. Variants that exhibit increased activity post-heat treatment are selected for further mutagenesis and screening cycles. Directed evolution has recently been used to select more stable phytase variants by introducing mutations that strengthen hydrophobic cores, elevate salt bridges, and improve hydrogen bonding networks [12].
3.2. Rational design to improve structural stability of phytase
Enzyme engineering by rational design is a fundamental and knowledge-based approach in which the structure of an enzyme is modified based on a thorough understanding of its catalytic mechanism and three-dimensional conformation [13]. Certain modifications to phytase have been successfully achieved by rational design (Figure 3).
High-resolution phytase structures help identify important residues for catalysis, stability, and substrate binding. Important amino acids in flexible surface loops, are often targeted for stabilization. Site-directed mutagenesis introduces specific mutations based on structural insights, using primers to insert changes via PCR. Cysteine residues may be added to form disulfide bridges, enhancing structural rigidity. The mutant phytase is expressed, purified, and tested under stress conditions to assess stability and activity. Test results guide for further improvements. Rational design has proven effective in enhancing phytase thermostability by reinforcing disulfide bonds and optimizing core hydrophobic interactions [12, 14, 17]. The applied rational design on E. coli phytase using molecular dynamics simulations and bioinformatics analyses were applied to identify a mutation (S392F) [14].
The mutant shows a significant increase in thermostability. It has increased activity at higher temperatures compared to the wild-type enzyme. Mutant displays two times higher thermostability at 70°C, and at 80°C and 90°C, it showed increased activity of 74% and 78.4%, respectively.
Secondary structure modifications further improve phytase stability. Flexible loops are critical for substrate access but vulnerable to thermal denaturation. They are stabilized by introducing proline residues to restrict conformational flexibility. A novel KeySIDE technique was used to improve the thermostability of Yersinia mollaretii phytase (Ymphytase) [16]. Nine important spots were found by combining directed evolution with iterative substitution analysis, and the optimum mutant, M6 (T77K, Q154H, G187S, K289Q), was developed. This mutant exhibited a significant increase in residual activity from 35% (wild-type) to 89% after 20 min at 58°C, with a 3°C rise in melting temperature and no loss of specific activity. Substitutions in loops close to helices B, F, and K (T77K, G187S, K289E/Q) decreased loop flexibility, based on molecular dynamics simulations. This caused by stronger hydrogen bonding networks (G187S, K289E/Q) and a salt bridge (T77K), which stabilized secondary structures and improved thermostability. Melting point bacterial phytase was raised by 7.5°C by strengthening its α-helices and β-sheets by improved hydrogen bonding or hydrophobic packing, strengthening the structural scaffold of the enzyme.
Evolution-guided rational design utilized to enhance thermostability of Yersinia intermedia phytase APPA [17]. They found high-frequency N-X-T/S motifs as potential spots for N-glycosylation modifications by examining 5569 homologous sequences from the NCBI database, with a focus on areas that are exposed to the surface. The optimum mutant M14 was created by combining mutations. It found to have enhanced kinetic stability and preserved wild-type catalytic efficiency. Its half-life (t1/2) increased from 3.32 min at 65°C to 25 min at 100°C, and retained 75% activity after 10 min at 100°C. When compared to wild-type APPA, in vitro digestibility of M14 increased by 4.5 times due to its improved kinetic stability and strong refolding ability, but its thermodynamic stability remained unchanged.
3.3. Computational design for predictive phytase engineering
Enzyme forward engineering uses computational tools to predict mutations that enhance activity and stability. Molecular modeling, dynamics (MD) simulations, and machine learning provide atomic-level insights. In phytase, these methods predict mutation effects and identify unstable regions under heat or acidity. Computer tools like Rosetta, AlphaFold, and FoldX design mutations to stabilize structure and improve binding [18]. Rosetta generates mutations that improve hydrophobic packing or stabilize loops using energy-based scoring. AlphaFold2 guides mutation selection by predicting 3D structures with high accuracy. FoldX identifies stabilizing modifications by estimating changes in free energy. Molecular dynamics simulations conducted using force fields i.e., AMBER ff14SB, CHARMM36, or OPLS-AA and software i.e., GROMACS, AMBER, or NAMD. They identify areas for stabilization by disulphide bond formation or electrostatic optimization by simulating phytase dynamics under thermal or acidic stress. Comprehensive enzyme sequence databases have been used to train machine learning systems like DeepMutate and ProteinMPNN, which accurately predict the effects of mutations and provide experimental confirmation. A major advantage of computational design is the ability to analyze a vast mutational space in silico, significantly reducing the number of studies required. Computational design detects modifications that improve thermostability by forming disulfide bonds, optimizing electrostatic interactions, or increasing hydrophobic packing. Experimental verification of the above theories allows the results to be fed back into computer models to enhance designs further [18, 19].
3.4. Semi-rational design integrating structural and sequence analysis
Semi-rational design intermediate directed evolution and rational design, requires knowledge of enzyme structure–function relationships. Important residues affecting stability, activity, or specificity are identified using biochemical data, structural analysis, and sequence alignment [13]. Phytases from fungi, bacteria, and plants share catalytic motifs but vary in heat and pH tolerance. Sequence alignment pinpoints residues linked to these traits, which are then modified via site-directed mutagenesis. Sequence analysis, bioinformatics, and evolutionary data are used in semi-rational design to identify potential mutation sites. This method creates targeted mutant libraries by utilizing techniques like multiple sequence alignments or directed evolution to target areas of interest, such as conserved motifs or variable loops. In comparison to directed evolution, this method allows the examination of a wide range of mutational regions with less experimental effort [13].
3.5. De novo design of synthetic phytase enzymes
De novo design creates novel enzymes with desired properties using computational methods, rather than modifying existing ones. It combines knowledge of protein folding, catalysis, and structural biology. In phytase engineering, it aims to build stable protein scaffolds with optimized active sites for phytate hydrolysis. The process uses modeling, energy minimization, and molecular dynamics to ensure structural stability and activity. Once a suitable design is achieved, the gene is synthesized and expressed in a host organism for validation [19, 20].
An automated workflow, GRACE (Generative Redesign in Artificial Computational Enzymology) is developed for reformation and creation of the de novo enzymes. It has integrated RFdiffusion (structure generation), ProteinMPNN (sequence interpretation), CLEAN (enzyme classification), solubility prediction, and molecular dynamics [19]. Two carbonic anhydrase-like enzymes, dCA12_2 and dCA23_1, were selected across 10000 protein options. Both demonstrated significant substrate-active site interactions, high solubility, and 400 WAU.mL-1 of enzymatic activity. GRACE significantly simplifies the design of enzymes. The alignment of 13 fungal phytase sequences was used to de novo create a consensus phytase in order to improve thermostability. The consensus enzyme showed a 15–22 °C higher unfolding temperature than any of its parent enzymes while retaining normal catalytic activity. According to a structural comparison with Aspergillus niger phytase, consensus residues significantly improved stability. Mutational study revealed that even those consensus amino acids that were assumed to be destabilizing instead improved resistance to heat. This work demonstrates an evident and unfamiliar connection between improved protein stability in fungal phytases and sequence conservation [20].
3.6. Glycosylation engineering for enhanced phytase stability
Fusion proteins are proteins formed by combining two or more genes that formerly coded for individual proteins. Domain swapping is a process for exchanging domains between related proteins to create hybrids with specific characteristics [17]. These domains are genetically combined or exchanged between proteins. Develop and assess the resultant mutant proteins to ensure they possess the required properties. The fusion protein formed by integrating a thermostable domain with a phytase enzyme improves the overall stability and performance of enzyme at a commercial scale. Directed evolution method have been studied to create phytase variants with increased thermostability [21]. This involved evaluating homologous sequences to find appropriate sites for introducing N-glycosylation variations, followed by site-directed mutagenesis. The engineered phytase variants found to have enhanced thermostability and high activity at boiling temperatures. The introduction of N-glycosylation sites stabilized the enzyme structure under extreme conditions.
Recent studies have demonstrated the benefits of introducing or optimizing N-glycosylation sites in phytases. E. coli phytase (AppA) involved introducing two N-glycosylation sites (Q258N and Q349N). After exposure to 85°C for 10 minutes, resulted in a mutant with a specific activity of 3137 mg.U-1 and over 40% enhanced thermostability. The melting temperature (Tₘ) increased by 4–5°C compared to the wild type, without affecting catalytic efficiency [22]. Glycosylation engineering of phytase from Cronobacter turicensis expressed in Pichia pastoris revealed that specific N-glycosylation sites (N136, N171, and N202) significantly influenced enzyme stability. Removal of the glycan at N202, located in a flexible region, led to a decrease in resistance to pepsin and trypsin by 73% and 87%, respectively. The glycosylated enzyme exhibited strong resistance to proteolytic degradation and increased thermostability [23]. An evolution-guided design approach analyzed 5569 homologous phytase sequences to identify 25 candidate sites for N-glycosylation. Thirteen of these sites were located on solvent-accessible surfaces and were selected for engineering. The introduction of N-glycosylation motifs at these positions enhanced the thermostability of enzyme. This demonstrates the effectiveness of combining evolutionary analysis with structural modelling in glycosylation engineering [17].
3.7. Advancements in phytase engineering
Recent advances in phytase engineering have considerably improved the thermostability and activity of enzymes. This expanded its application in a range of food processing conditions. Phytases have significance for increasing nutrient bioavailability in plant-based foods and feeds. However, their poor stability at high temperatures has limited their industrial applications. A comprehensive study reviewed the advances achieved by producing thermostable phytases over the last seven years using genetic alterations and immobilization techniques. The study indicated that protein engineering techniques have successfully improved the thermostability and activity of phytases. Immobilization on different supports has resulted in 50-60% activity retention at temperatures above 50°C, demonstrating the potential of these techniques in industrial applications [24].
A targeted examination was carried out to change the phytase YiAPPA by mutating protein surface residues [25]. They utilized site-directed mutagenesis to create single-site mutants that substituted specific amino acids on the surface of enzyme. This technique produced mutants with higher thermostability and activity, increasing their potential for use in the food industry. Furthermore, the wild-type YiAPPA showed a half-life of about 30 min at 55°C, which significantly improved in the engineered variants. E. coli phytase thermostability is enhanced using error-prone polymerase chain reaction (epPCR). Directed evolution is effective in improving the thermostability of phytases. Variants were first expressed in E. coli BL21 and simultaneously in Pichia pastoris for screening and characterization, respectively. Researchers developed phytase variants capable of functioning at the high temperatures found in industrial processes by applying multiple stages of mutation and selection. Mutants had higher residual activity and catalytic efficiency, ranging from 9.6% to 12.2% and 79.8% to 92.6%, respectively. It was determined that the C77-C108 disulfide link in E. coli phytase was crucial for stability [26]. However, balancing higher temperature stability with the preservation-specific enzyme activity remains challenging. Disulfide bond engineering is another potential technique. Researchers improved the thermostability of phytase by inserting more disulfide bonds. Modifications to the AppA phytase have resulted in increased stability, making it more suitable for industrial applications requiring high-temperature processes [33]. Modifications to the AppA phytase have resulted in increased stability, which makes it more suitable for industrial applications requiring high-temperature processes [33]. In food industry processes such as baking, extrusion, and fermentation, phytases must withstand temperatures exceeding 50–80 °C and operate effectively within a pH range of 4.0–6.5. Engineered variants that retain 50–75% activity under these conditions demonstrated significant commercial potential. This is helpful in flour fortification and bread-making where phytate hydrolysis is essential for mineral bioavailability [24, 33, 36].
3.8. Limitations of current engineering techniques
Each phytase engineering technique offers distinct advantages but also presents specific limitations. Directed evolution is an effective at generating improved variants. However, it relies on random mutagenesis, requires the screening of vast mutant libraries. This process is labor-intensive, time-consuming, and often inefficient, with a risk of losing catalytic efficiency or substrate specificity. Rational design provides a more targeted approach but depends heavily on detailed structural knowledge and high-resolution enzyme models. Its success is limited by the accuracy of structural predictions and may not always result in significant functional improvements. Computational design involves high computational costs and requires extensive knowledge in molecular modeling and simulations. Moreover, in silico predictions may not always translate accurately to real-world enzyme properties, and experimental validation remains essential. Semi-rational design offers a balance between random and structure-based methods but may miss important mutational hotspots and still requires substantial experimental screening. De novo design is the most advanced method, but it is technically complex and prone to challenges i.e., misfolding, low catalytic activity, and unpredictable stability under industrial conditions. Glycosylation engineering enhances enzyme stability through post-translational modification. It is limited by host-specific glycosylation patterns, potential interference with the active site, and increased production complexity. Despite significant progress in these areas, common industrial limitations i.e., thermal instability, high production costs, inefficient high-throughput screening, and a lack of robust enzymes that function across a wide range of pH and temperature conditions remain major challenges. Overcoming these challenges will require integrated, multidisciplinary approaches that combine synthetic biology, machine learning, and cost-effective production methods.
- Applications of engineered phytase in the food industry
Engineered phytases have drawn a lot of attention due to their capacity to hydrolyze phytic acid, a common antinutrient found in grains and legumes, improving nutritional profiles and increasing mineral bioavailability. Production of phytases with improved properties, diversified their application in food processing as a result of development and advancement in genetic engineering.
4.1. Phytase application in baked products for mineral bioavailability
Engineered phytases in bread-making are gaining attention for improving the nutritional value of whole-grain products. Phytic acid in grains, legumes, and seeds, binds minerals like calcium, zinc, and iron, forming insoluble complexes that reduce their bioavailability. This can lead to nutrient deficiencies in plant-based diets. Phytases help break down phytic acid, either through added microbial enzymes or by activating natural cereal phytases during dough fermentation. Optimizing dough pH enhances phytase activity, effectively reducing phytic acid content. [9] summarized human intervention studies concluded that majority of researches evaluating the exogenous phytase efficacy or food dephytinization resulted in enhanced bioavailability of zinc and iron. The degree of phytic acid breakdown is determined by the bread-making method used. Research comparing several wheat varieties and bread-making methods has demonstrated that the baking method has a significant effect on the final phytic acid level than the wheat variety selected. Long yeast fermentation methods were more efficient in lowering phytic acid levels and improving bioavailability of mineral.
A novel protein tyrosine phosphatase-like phytase (PhyLf) was identified from Limosi lactobacillus fermentum NKN51. The recombinant enzyme exhibited a specific activity of 174.5 U.mg-1, with an optimal temperature, pH, and ionic strength of 60°C, 5.0 and 100 mM, respectively. Its shown ability to enhance mineral bioavailability under in vitro gastrointestinal conditions in finger millet and durum wheat. This highlighted its potential for improving the nutritional value of cereals and animal feed [34]. Two directed‐mutagenesis S. cerevisiae strains (YD80, BY80) and a naturally high‐phytase Pichia kudriavzevii TY13 were employed in wheat–cassava–sorghum composite bread. They achieved up to 99 % reduction in phytic acid and phytate:iron and phytate:zinc molar ratios well below critical thresholds for human absorption (1:1 and 15:1, respectively) [35]. Pre‐incubation of the yeasts at 30 °C for 1 h and supplementation with yeast extracts further enhanced phytase biosynthesis, leading to the lowest measured phytic acid content of 0.08 µmol/g in the final bread. [36] added a commercial A. niger phytase was added to whole‐wheat bread dough, observing that although proofing degraded phytic acid by up to 49.4 %, complete hydrolysis was affected due to bran particle size limiting enzyme access [36]. This resulted in residual phytate levels of 3.74 mg.g-1 dry matter in baked loaves. This highlighted the need for enzyme engineering or formulation to improve substrate accessibility during baking.
In Sangak and Barbari breads, exogenous A. niger phytase addition significantly increased in vitro iron, zinc, and manganese availability without affecting copper levels, demonstrating that targeted phytase application can selectively enhance key mineral bio accessibility in traditional flatbreads. The study reported up to a twofold increase in iron and zinc solubility post‐baking, underscoring potential for regional dietary interventions [37]. Incorporating phytase into baked goods can reduce phytic acid content, enhancing mineral bioavailability. However, the high temperatures involved in baking can inactivate phytase, necessitating the use of thermostable variants. Studies have demonstrated that certain phytase enzymes retain activity after baking, making them suitable for fortifying bread and other baked products.
4.2. Role of phytase in fermentation for nutrient enhancement
In fermented foods, phytase affects the fermentation and affect flavor development. Engineered phytases are introduced either by adding purified enzymes or using phytase-producing microbes. The microbial approach involves selecting or genetically modifying strains i.e., lactic acid bacteria (LAB), to produce the enzyme during fermentation. Some of LAB naturally possess phytase activity. They reduce phytic acid during food processing and gastrointestinal digestion, increasing mineral bioavailability and the nutritional profile of the final product [38]. The second approach to reducing phytic acid is the direct addition of purified phytases. Advances in genetic engineering have produced phytases with improved temperature stability, pH tolerance, and substrate specificity, suitable for various food processing conditions. Adding microbial phytases to foods effectively lowers phytic acid content, enhancing mineral bioavailability and overall nutritional quality.
The scald fermentation of rye (12–48 h) with selected lactic acid bacteria starters was investigated finding that prolonged fermentation reduced phytate content by over 60 % and improved nutritional profile of rye bread, including enhanced sensory qualities [39]. The study demonstrates that process‐driven activation of endogenous and microbial phytases during fermentation can rival direct enzyme additions in lowering phytic acid. During tempeh production from soybeans inoculated with Rhizopus oligosporus, endogenous fungal phytase halves phytic acid in the first 24 h, and extended storage (72 h at 5 °C or 30 °C) plus deep-fat frying further reduce residual phytate to below 10 % of the original level [40]. Koji molds found to be differ in phytase thermostability during soy sauce brewing. After 30 days, A. sojae–fermented mash shows undetectable phytic acid, whereas A. oryzae retains measurable levels, attributed to the lower heat resilience of its phytase enzyme [41]. An extracellular phytase from Lactobacillus sakei (kimchi‐derived) has an optimum pH of 5.5 and hydrolyzes approximately 60 % of phytic acid in model fermentations within 24 h, improving availability of iron and zinc in the final product [42].
4.3. Development of functional foods and supplements
Incorporating engineered phytases into functional foods and supplements offers a promising approach to prevent mineral deficiencies in populations reliant on phytate-rich staples. These enzymes enhance mineral absorption by breaking down phytic acid into inositol and inorganic phosphate. Microbial phytases from fungi and bacteria are effective across a wide range of pH and temperatures, making them suitable for diverse food applications. This approach is beneficial in developing regions where diets are heavily based on cereals and legumes with high phytic acid content. Phytate content in the milk was reduced using the immobilized phytase on starch agar beads [43]. Processed soymilk serves as major ingredient in baby nutrition. Reduced phytate levels increase the bioavailability of minerals and proteins in infants.
Fermentation, soaking, and germination activate natural phytases or enhance microbial phytase action. These reduce the phytic acid content and enhance bioavailability of micronutrients. These methods improve staple food nutrition in regions with mineral deficiencies. Incorporating engineered phytases in functional foods and supplements addresses these deficiencies with improved food processing. Breaking down phytic acid enhances the quality and yield of products like bread, plant protein isolates, and cereal bran. This demonstrates the role of phytase in improving nutrition and functionality.
Certain strains of Bifidobacteria i.e., B. pseudocatenulatum and B. longum, have been utilized in sourdough fermentation to produce phytase in situ. This fermentation process significantly reduces phytic acid levels in whole rye-wheat mixed bread and increased dialyzable iron content by 2.3- to 5.6-fold. These findings suggest that incorporating phytase-producing probiotics into bread making can enhance mineral bioavailability without compromising sensory qualities [44]. Tolerase® P is a phytase developed by DSM-Firmenich. It has demonstrated efficacy in improving mineral absorption from plant-based foods. Clinical studies have shown that this enzyme can increase iron absorption by up to 11.6 times and zinc absorption by up to 2 times. Its stability across a broad pH range (2.5–5.5) and compatibility with various food matrices make it suitable for incorporation into flour fortification, cereals, and dietary supplements [45].
- Concluding remarks and future perspectives
Phytase enzyme engineering methods have achieved significant progress in terms of thermostability, catalytic efficiency, and resistance to challenging industrial environments. The application of modern molecular techniques such as directed evolution, rational design, and computational modelling results in the development of more effective phytase variants with enhanced functionality. These developments expand the application of phytase in food processing, especially in increasing the nutritional value by breaking down phytic acid and increasing mineral bioavailability of plant-based foods. Incorporation of engineered phytase into the food and feed industries has the potential to address major challenges i.e., mineral deficiencies and sustainable food production. Future research should focus on optimizing large-scale phytase production, improving enzyme stability under a range of processing conditions, and promoting its use in functional foods and probiotic formulations. As enzyme engineering improves, phytase will play an increasingly important role in enhancing food nutrition and reducing environmental phosphorus pollution, subsequently increasing human health and ecological sustainability.
Furthermore, the economic feasibility of using engineered phytases into large-scale food production remains a critical factor for industrial adoption. High costs associated with enzyme purification, stabilization, and immobilization limits widespread use. Therefore, future innovations must focus on cost-effective purification systems, scalable fermentation technologies, and reusable immobilization supports to enhance process economics. In parallel, regulatory approval presents significant hurdles, notably for genetically modified enzymes. Acceptance and clearances from regulatory bodies requires rigorous safety assessments and compliance with labeling regulations. Additionally, consumer acceptance of phytase-treated foods may vary due to perceptions surrounding genetic modification and enzyme-treated products. To ensure safety and successful market adoption of phytase-enhanced food products, transparent labeling, public education, and clinical evidence of nutritional benefits will be essential.
- Acknowledgments
This project was conducted using the resources of the Department of Food Process Engineering, National Institute of Technology, Rourkela, Odisha, India.
- Declaration of competing interest
The authors report no conflict of interest.
- Authors’ Contributions
Writing-original draft, C.M.; conceptualization, C.M.; methodology, C.M. and A.K.; visualization, C.M.; data curation, R.C.P.; supervision, R.C.R. and R.C.P.; project administration, R.C.R.; writing-review & editing, R.C.R. and R.C.P.; resources, A.K. and R.C.P.
- Using Artificial Intelligent Chatbots
No artificial intelligent chatbots has been used in any section of work.
- Phytase
- enzyme engineering
- directed evolution
- food processing
- phytic acid
- mineral bioavailability
How to Cite
References
Hussain SM, Hanif S, Sharif A, Bashir F, Iqbal HMN. Unrevealing the sources and catalytic functions of phytase with multipurpose characteristics. Catal Lett. 2021; 152(5): 1358-1371. https://doi.org/10.1007/s10562-021-03752-z
Gorissen SHM, Crombag JJR, Senden JMG, Waterval WaH, Bierau J, Verdijk LB, Van Loon LJC(2018). Protein content and amino acid composition of commercially available plant-based protein isolates. Amino Acids. 2018; 50(12): 1685-1695. https://doi.org/10.1007/s00726-018-2640-5
Liang Q, Yuan M, Xu L, Lio E, Zhang F, Mou H, Secundo F. Application of enzymes as a feed additive in aquaculture. Mar Life Sci Technol. 2022; 4(2): 208-221. https://doi.org/10.1007/s42995-022-00128-z
Hartwell CJ, Johnson CD, Posthuma RA. Are we asking the right questions? Predictive validity comparison of four structured interview question types. J Bus Res. 2019; 100: 122-129. https://doi.org/10.1016/j.jbusres.2019.03.026
Feizollahi E, Mirmahdi RS, Zoghi A, Zijlstra RT, Roopesh, Vasanthan T. Review of the beneficial and anti-nutritional qualities of phytic acid, and procedures for removing it from food products. Food Res Int. 2021; 143: 110284. https://doi.org/10.1016/j.foodres.2021.110284
Kumari A, Roy A. Enhancing micronutrient absorption through simultaneous fortification and phytic acid degradation. Food Sci Biotechnol. 2023; 32(9): 1235-1256. https://doi.org/10.1007/s10068-023-01255-8
Kumar V, Sinha AK, Makkar HPS, De Boeck G, Becker K. Phytate and phytase in fish nutrition. J Anim Physiol Anim Nutr. 2011; 96(3): 335-364. https://doi.org/10.1111/j.1439-0396.2011.01169.x
Kumar V, Singh D, Sangwan P, Gill PK. Management of Environmental Phosphorus Pollution Using Phytases: Current Challenges and Future Prospects, In: Springer eBooks. 2015; p.97-114. https://doi.org/10.1007/978-81-322-2123-4_7
Chondrou T, Adamidi N, Lygouras D, Hirota SA, Androutsos O, Svolos V. Dietary phytic acid, dephytinization, and phytase supplementation alter trace element bioavailability—A narrative review of human interventions. Nutrients. 2024; 16(23): 4069. https://doi.org/10.3390/nu16234069
Gunathunga C, Senanayake S, Jayasinghe MA, Brennan CS, Truong T, Marapana U, Chandrapala J. Germination effects on nutritional quality: A comprehensive review of selected cereals and pulses changes. J Food Compos Anal. 2024; 128: 106024. https://doi.org/10.1016/j.jfca.2024.106024
Raboy V, Gibson RS, Bailey KB, King JC. Comparison of four methods for phytate analysis in plant-based foods. J Food Compos Anal. 2020; 90: 103481. https://doi.org/10.1016/j.jfca.2020.103481
Wang Y, Xue P, Cao M, Yu T, Lane ST, Zhao H. Directed Evolution: Methodologies and applications. Chem Rev. 2021; 121(20): 12384-12444. https://doi.org/10.1021/acs.chemrev.1c00260
Song Z, Zhang Q, Wu W, Pu Z, Yu H. Rational design of enzyme activity and enantioselectivity. Front Bioeng Biotechnol. 2023; 11. https://doi.org/10.3389/fbioe.2023.1129149
Fakhravar A, Hesampour A. Rational design-based engineering of a thermostable phytase by site-directed mutagenesis. Mol Biol Rep. 2018; 45(6): 2053-2061. https://doi.org/10.1007/s11033-018-4362-x
Daletos G, Stephanopoulos G. Protein engineering strategies for microbial production of isoprenoids. Metab Eng Commun. 2020; 11:e00129. https://doi.org/10.1016/j.mec.2020.e00129
Shivange AV, Roccatano D, Schwaneberg U. Iterative key-residues interrogation of a phytase with thermostability increasing substitutions identified in directed evolution. Appl Microbiol Biotechnol. 2015; 100(1): 227-242. https://doi.org/10.1007/s00253-015-6959-5
Wang Q, Liu X, Tian J, Wang Y, Zhang H, Wang Y, Luo H, Yao B, Huang H, Tu T. Enhancing the thermostability of phytase to boiling point by Evolution-Guided design. Appl Environ Microbiol. 2022; 88(11): e00506-22. https://doi.org/10.1128/aem.00506-22
Peccati F, Alunno-Rufini S, Jiménez-Osés G. Accurate Prediction of Enzyme Thermostabilization with Rosetta Using AlphaFold Ensembles. J Chem Inf Model. 2023; 63(3): 898-909. https://doi.org/10.1021/acs.jcim.2c01083
Hu R, Yu C, Ng I. GRACE: Generative Redesign in Artificial Computational Enzymology. ACS Synth Biol. 2024. https://doi.org/10.1021/acssynbio.4c00624
Lehmann M, Kostrewa D, Wyss M, Brugger R, D’Arcy A, Pasamontes L, Van Loon AP. From DNA sequence to improved functionality: using protein sequence comparisons to rapidly design a thermostable consensus phytase. Protein Eng Des Sel. 2000; 13(1):49-57. https://doi.org/10.1093/protein/13.1.49
Wang Q, Wang T, Zhang R, Yang S, McFarland KS, Chung C, Jia H, Wang L, Cipollo JF, Betenbaugh M.J. The interplay of protein engineering and glycoengineering to fine‐tune antibody glycosylation and its impact on effector functions. Biotechnol Bioeng. 2021; 119(1): 102-117. https://doi.org/10.1002/bit.27953
Yao M, Wang X, Wang W, Fu Y, Liang A. Improving the thermostability of Escherichia coli phytase, appA, by enhancement of glycosylation. Biotechnol Lett. 2013; 35(10): 1669-1676. https://doi.org/10.1007/s10529-013-1255-x
Gordeeva TL, Borshchevskaya LN, Sineoky SP. Biochemical characterisation of glycosylated and deglycosylated forms of phytase from Cronobacter turicensis expressed in Pichia pastoris. Enzyme Microb Technol. 2022; 162: 110136. https://doi.org/10.1016/j.enzmictec.2022.110136
Venkataraman S, Karthikanath PR, Gokul CS, Adhithya M, Vaishnavi VK, Rajendran DS, Vaidyanathan VK, Natarajan R, Balakumaran PA, Kumar VV. Recent advances in phytase thermostability engineering towards potential application in the food and feed sectors. Food Sci Biotechnol. 2024; 34(1): 1-18. https://doi.org/10.1007/s10068-024-01690-1
Zeng J, Guo J, Yuan L. Engineering of the phytase YIAPPA to improve thermostability and activity and its application potential in dephytinization of food ingredients. J Microbiol Biotechnol. 2024; 34(8): 1660-1670. https://doi.org/10.4014/jmb.2403.03031
Xing H, Wang P, Yan X, Yang Y, Li X, Liu R, Zhou Z. Thermostability enhancement of Escherichia coli phytase by error-prone polymerase chain reaction (epPCR) and site-directed mutagenesis. Front Bioeng Biotechnol. 2023; 11. https://doi.org/10.3389/fbioe.2023.1167530
Tu T, Wang Q, Dong R, Liu X, Penttinen L, Hakulinen N, Tian J, Zhang W, Wang Y, Luo H, Yao B, Huang H. Achieving thermostability of a phytase with resistance up to 100°C. J Biol Chem. 2024: 107992. https://doi.org/10.1016/j.jbc.2024.107992
Niu C, Yang P, Luo H, Huang H, Wang Y, Yao B. Engineering the residual side chains of HAP phytases to improve their pepsin resistance and catalytic efficiency. Sci Rep. 2017; 7(1). https://doi.org/10.1038/srep42133
Muñoz-Muñoz PLA, Terán-Ramírez C, Mares-Alejandre RE, Márquez-González AB, Madero-Ayala PA, Meléndez-López SG, Ramos-Ibarra MA. Surface engineering of Escherichia coli to display its phytase (AppA) and functional analysis of enzyme activities. Curr Issues Mol Biol. 2024; 46(4): 3424-3437. https://doi.org/10.3390/cimb46040215
Bandari NM, Abootaleb M, Nikokar I, Karimli M. Biologically engineered probiotic supplement production containing phytase enzyme for livestock, poultry, and aquaculture consumption. J Basic Appl Zool. 2024; 85(1). https://doi.org/10.1186/s41936-024-00361-1
Liao Y, Zeng M, Wu Z, Chen H, Wang H, Wu Q, Shan Z, Han X. Improving phytase enzyme activity in a recombinant phyA mutant phytase from Aspergillus niger N25 by error-prone PCR. Appl Biochem Biotechnol. 2011; 166(3): 549-562. https://doi.org/10.1007/s12010-011-9447-0
Zhao Y, Zhu L, Lin C, Shen Z, Xu C. Transgenic soybean expressing a thermostable phytase as substitution for feed additive phytase. Sci Rep. 2019; 9(1). https://doi.org/10.1038/s41598-019-51033-y
Navone L, Vogl T, Luangthongkam P, Blinco J, Luna-Flores CH, Chen X, Von Hellens J, Mahler S, Speight R. Disulfide bond engineering of AppA phytase for increased thermostability requires co-expression of protein disulfide isomerase in Pichia pastoris. Biotechnol Biofuels. 2021; 14(1). https://doi.org/10.1186/s13068-021-01936-8
Sharma R, Kumar P, Kaushal V, Das R, Navani NK. A novel protein tyrosine phosphatase like phytase from Lactobacillus fermentum NKN51: Cloning, characterization and application in mineral release for food technology applications. Bioresour. Technol. 2017; 249: 1000-1008. https://doi.org/10.1016/j.biortech.2017.10.106
Vilanculos SL, Svanberg U, Andlid T. Phytate degradation in composite wheat/cassava/sorghum bread: Effects of phytase‐secreting yeasts and addition of yeast extracts. Food Sci Nutr. 2023; 12(1): 216-226. https://doi.org/10.1002/fsn3.3754
Haros M, Rosell CM, Benedito C. Fungal phytase as a potential breadmaking additive. Eur Food Res Technol. 2001; 213(4–5): 317-322. https://doi.org/10.1007/s002170100396
Shockravi S, Almgren A, Carlsson N, Sandberg A. Dephytinisation of Sangak and Barbari bread made from different extraction rate flours increases iron and zinc bioaccessibility in Caco‐2 cells. Int J Food Sci Technol. 2012; 47(11): 2252-2258. https://doi.org/10.1111/j.1365-2621.2012.03095.x
Sharma N, Angural S, Rana M, Puri N, Kondepudi KK, Gupta N. Phytase-producing lactic acid bacteria: Cell factories for enhancing micronutrient bioavailability of phytate-rich foods. Trends Food Sci Technol. 2019; 96: 1-12. https://doi.org/10.1016/j.tifs.2019.12.001
Murniece R, Reidzane S, Radenkovs V, Straumite E, Keke A, Kobrin E, Klava D. Scald fermentation time as a factor determining the nutritional and sensory quality of rye bread. Foods. 2025; 14(6): 979. https://doi.org/10.3390/foods14060979
Duliński R, Stodolak B, Byczyński Ł, Poreda A, Starzyńska-Janiszewska A, Żyła K. Solid-state fermentation reduces phytic acid level, improves the profile of myo-inositol phosphates and enhances the availability of selected minerals in flaxseed oil cake. Food Technol Biotechnol. 2017; 55(3). https://doi.org/10.17113/ftb.55.03.17.4981
Ito K, Matsuyama A. Koji molds for Japanese soy sauce brewing: Characteristics and key enzymes. J Fungi. 2021; 7(8): 658. https://doi.org/10.3390/jof7080658
Park S, Yang S, Lee J, Kang M. Selection of phytate-degrading lactic acid bacteria from kimchi and reaction properties in brown rice. J Korean Soc Food Sci Nutr. 2013; 42(4): 627-632.
Ushasree MV, Shyam K, Vidya J, Pandey A. Microbial phytase: Impact of advances in genetic engineering in revolutionizing its properties and applications. Bioresour Technol. 2017; 245: 1790-1799. https://doi.org/10.1016/j.biortech.2017.05.060
Chen L, Tian F, Su Z. Phosphorus nutrition and health: Utilization of phytase-producing bifidobacteria in food industry. In: InTech eBooks. 2016. https://doi.org/10.5772/63644
dsm-firmenich. Tolerase® P: Breakthrough solution for better mineral absorption. 2025. https://www.dsm-firmenich.com/en/businesses/health-nutrition-care/news/talking-nutrition/how-phytase-breaks-through-absorption-barriers.html
- Abstract Viewed: 980 times
- pdf Downloaded: 328 times